

Comparative Proteomic Analysis of Desulfotomaculum reducens MI-1: Insights into the Metabolic Versatility of a Gram-Positive Sulfate- and Metal-Reducing Bacterium
- PMID: 26925055
- PMCID: PMC4759654
- DOI: 10.3389/fmicb.2016.00191
Comparative Proteomic Analysis of Desulfotomaculum reducens MI-1: Insights into the Metabolic Versatility of a Gram-Positive Sulfate- and Metal-Reducing Bacterium
Abstract
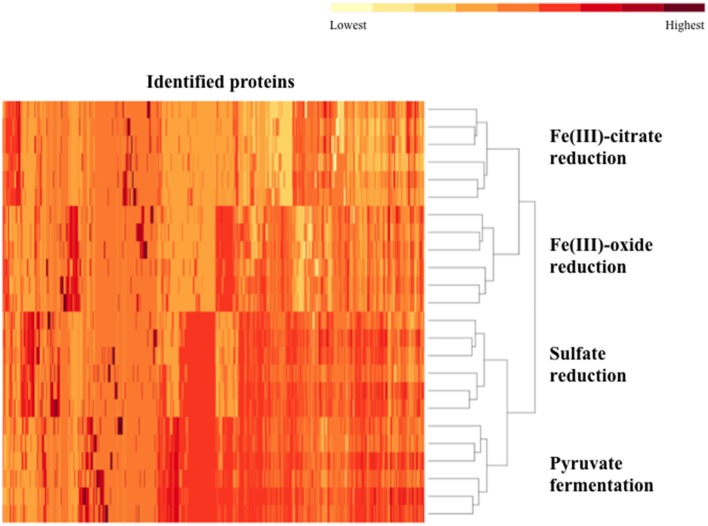
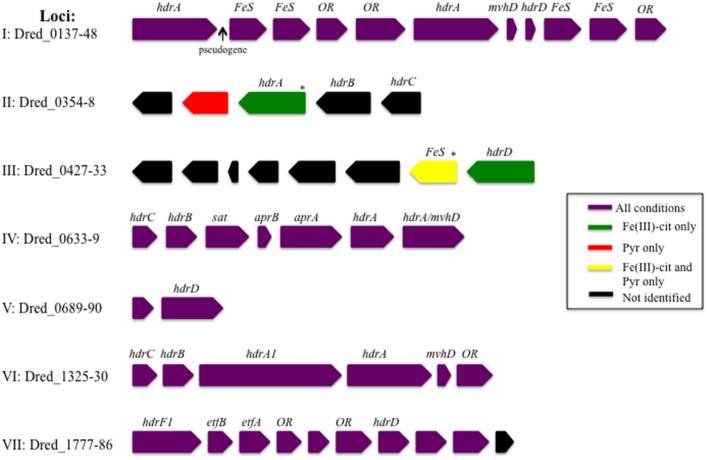
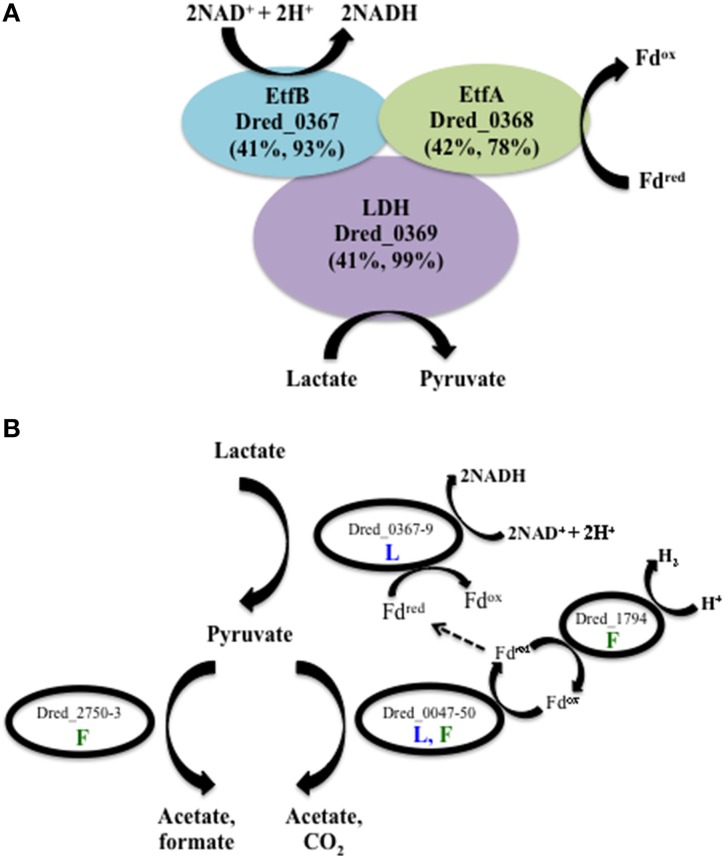
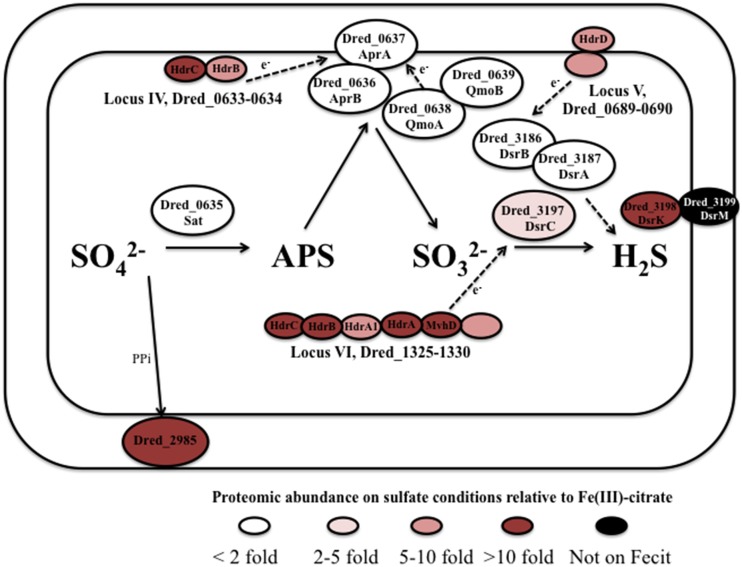
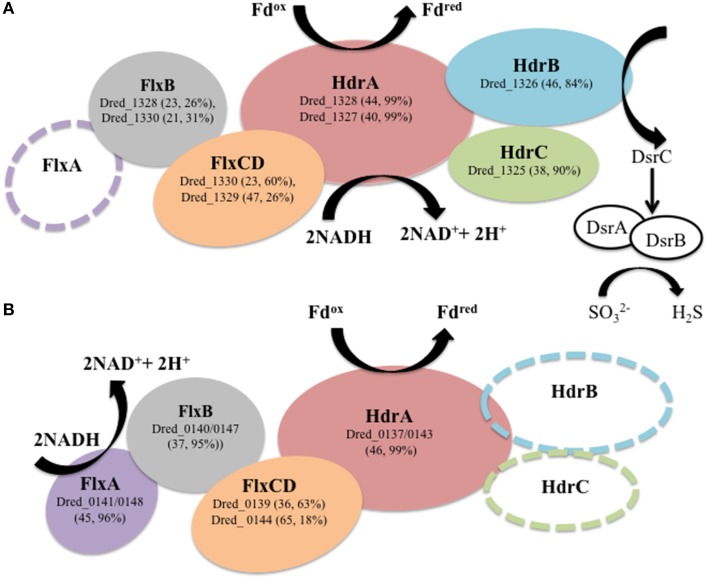
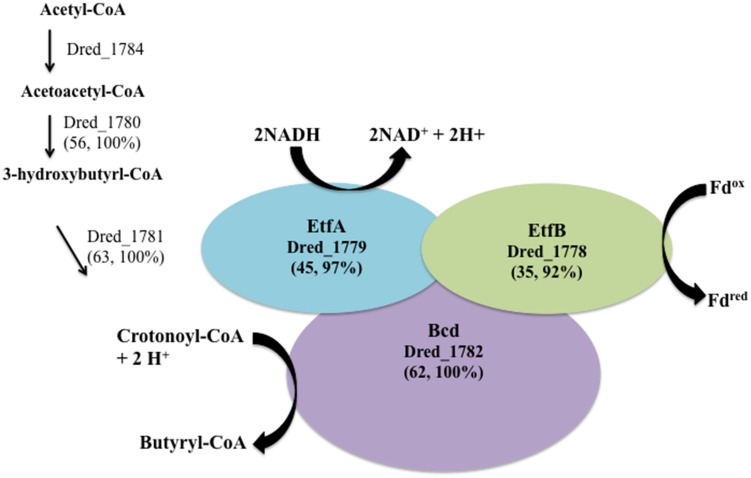
The proteomes of the metabolically versatile and poorly characterized Gram-positive bacterium Desulfotomaculum reducens MI-1 were compared across four cultivation conditions including sulfate reduction, soluble Fe(III) reduction, insoluble Fe(III) reduction, and pyruvate fermentation. Collectively across conditions, we observed at high confidence ~38% of genome-encoded proteins. Here, we focus on proteins that display significant differential abundance on conditions tested. To the best of our knowledge, this is the first full-proteome study focused on a Gram-positive organism cultivated either on sulfate or metal-reducing conditions. Several proteins with uncharacterized function encoded within heterodisulfide reductase (hdr)-containing loci were upregulated on either sulfate (Dred_0633-4, Dred_0689-90, and Dred_1325-30) or Fe(III)-citrate-reducing conditions (Dred_0432-3 and Dred_1778-84). Two of these hdr-containing loci display homology to recently described flavin-based electron bifurcation (FBEB) pathways (Dred_1325-30 and Dred_1778-84). Additionally, we propose that a cluster of proteins, which is homologous to a described FBEB lactate dehydrogenase (LDH) complex, is performing lactate oxidation in D. reducens (Dred_0367-9). Analysis of the putative sulfate reduction machinery in D. reducens revealed that most of these proteins are constitutively expressed across cultivation conditions tested. In addition, peptides from the single multiheme c-type cytochrome (MHC) in the genome were exclusively observed on the insoluble Fe(III) condition, suggesting that this MHC may play a role in reduction of insoluble metals.
Keywords: Desulfotomaculum reducens; Fe(III) reduction; comparative proteomic analysis; flavin-based electron bifurcation; gram-positive bacteria; heterodisulfide reductase; sulfate reduction.
Figures
References
-
- Barton L., Tomei-Torres F., Xu H., Zocco T. (2015). Metabolism of metals and metalloids by the sulfate-reducing bacteria, in Bacteria-Metal Interactions, ed Saffarini Cham D. (Cham: Springer International Publishing; ), 57–83.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials