

Review
doi: 10.1186/s13059-016-0908-1.
Evolution of plant genome architecture
Affiliations
- PMID: 26926526
- PMCID: PMC4772531
- DOI: 10.1186/s13059-016-0908-1
Item in Clipboard
Review
Evolution of plant genome architecture
Genome Biol.
.
Abstract
We have witnessed an explosion in our understanding of the evolution and structure of plant genomes in recent years. Here, we highlight three important emergent realizations: (1) that the evolutionary history of all plant genomes contains multiple, cyclical episodes of whole-genome doubling that were followed by myriad fractionation processes; (2) that the vast majority of the variation in genome size reflects the dynamics of proliferation and loss of lineage-specific transposable elements; and (3) that various classes of small RNAs help shape genomic architecture and function. We illustrate ways in which understanding these organism-level and molecular genetic processes can be used for crop plant improvement.
Figures
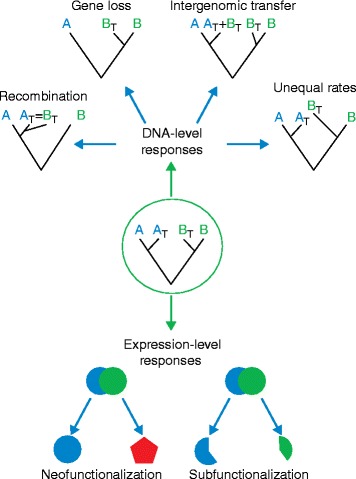
Processes and patterns of polyploidy in plants. Flowering plant genomes have an evolutionary history that includes multiple, lineage-specific, whole-genome doubling events. A model of a hypothetical allopolyploid genome derived from two progenitor diploid genomes (A and B) is shown in the figure. Hybridization and genome doubling set in motion short-term and long-term genomic processes, at the level of DNA sequence (top) and at the expression level (bottom). Young allopolyploids, which initially contain two genomes (A
T and B
T) inherited from the parental genome donors, often display homoeologous recombination (‘gene conversion’), mutational loss and/or silencing of duplicated genes, intergenomic spread of TEs and differential rates of molecular evolution, as well as many different forms of biased or altered gene expression, including biased homoeolog expression and expression level dominance (not shown), and expression subfunctionalization (shown as partitioning of ancestral blue and green expression domains, bottom left) and neofunctionalization (novel red expression domain, bottom right)
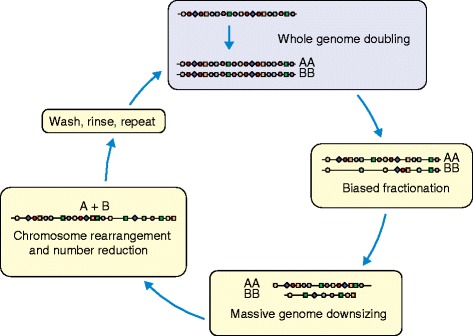
Episodic, cyclical episodes of polyploidy in plants. All plant lineages are paleopolyploid, differing only in the number of rounds of whole-genome doubling in their ancestry, and in the timing of these events. Long-term processes following whole-genome doubling include differential retention of duplicated genes (here, more circles than squares are lost), biased genome fractionation (shown as greater loss of the B genome), massive genome downsizing and reduction in the number of chromosomes
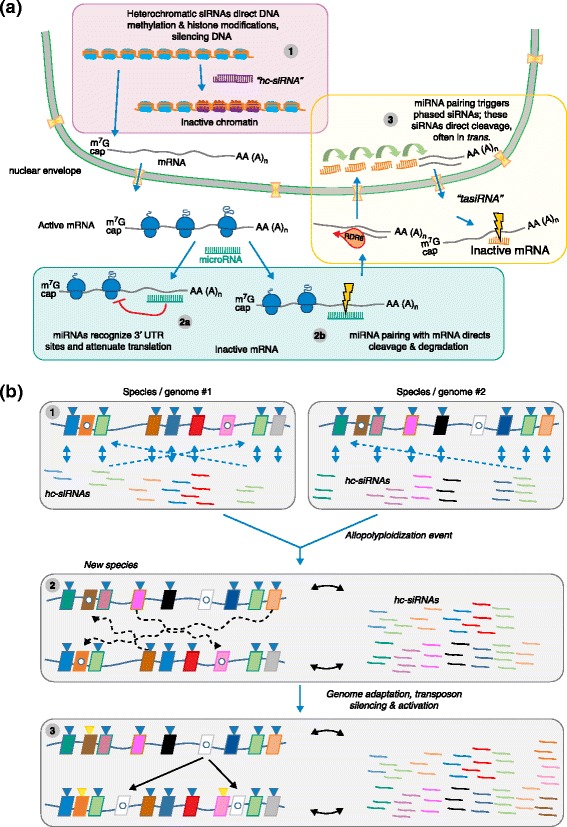
The role of small RNAs in plant genome evolution. a Plant genomes are rich sources of small RNAs, which are predominantly the products of three major pathways. (1) Heterochromatic siRNAs (hc-siRNAs) are 24-nucleotide products of the activity of the plant-specific Pol IV and Pol V pathways; hc-siRNAs derive from heterochromatic regions and target those regions for reinforcement of silencing chromatin marks. (2) MicroRNAs (miRNAs) are ~21-nucleotide or 22-nucleotide products of processed long noncoding mRNAs that function to suppress target mRNAs either by (2a) blocking translation, or (2b) directing Argonaute-mediated slicing of targets. Plant miRNAs typically function by directing slicing. (3) Some miRNAs, typically 22 nucleotides long, trigger the production of secondary siRNAs, which are products of DCL4 and are 21 nucleotide long, from their target mRNAs. This pathway requires RDR6, and some resulting siRNAs can act in trans to slice additional targets; hence their name of trans-acting siRNAs (tasiRNAs). b hc-siRNAs are typically derived from TEs, the predominant component of inactive chromatin in plant genomes. Transposons (colored parallelograms in 1–3) can be silenced (blue triangles) as a result of RNA-directed DNA methylation mediated by hc-siRNAs derived from those elements (thin blue arrows). Some transposons can escape DNA methylation and silencing (white dots), to later remobilize. Transposons can be additionally silenced by hc-siRNAs functioning in trans from related TEs (dashed lines). After allopolyploid individuals form (2), the genomic context changes for TEs from the progenitor genomes, and these elements can be silenced by hc-sRNAs derived from sequence-similar TEs residing in the added genome (dashed, curvy lines). After this period of adaptation (3), some TEs can be newly silenced (gold triangles), whereas a few might remain unsilenced and active, and might amplify into the added genome (thin black arrows). UTR untranslated region
References
-
- Grant V. Plant speciation. New York: Columbia; 1981.
-
- Stebbins GL. Chromosomal evolution in higher plants. London: Edward Arnold; 1971.
-
- Ramsey J, Schemske DW. Pathways, mechanisms, and rates of polyploid formation in flowering plants. Annu Rev Ecol Syst. 1998;29:467–501. doi: 10.1146/annurev.ecolsys.29.1.467. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
