

A few of our favorite things: Pairing, the bouquet, crossover interference and evolution of meiosis
- PMID: 26927691
- PMCID: PMC4867269
- DOI: 10.1016/j.semcdb.2016.02.024
A few of our favorite things: Pairing, the bouquet, crossover interference and evolution of meiosis
Abstract
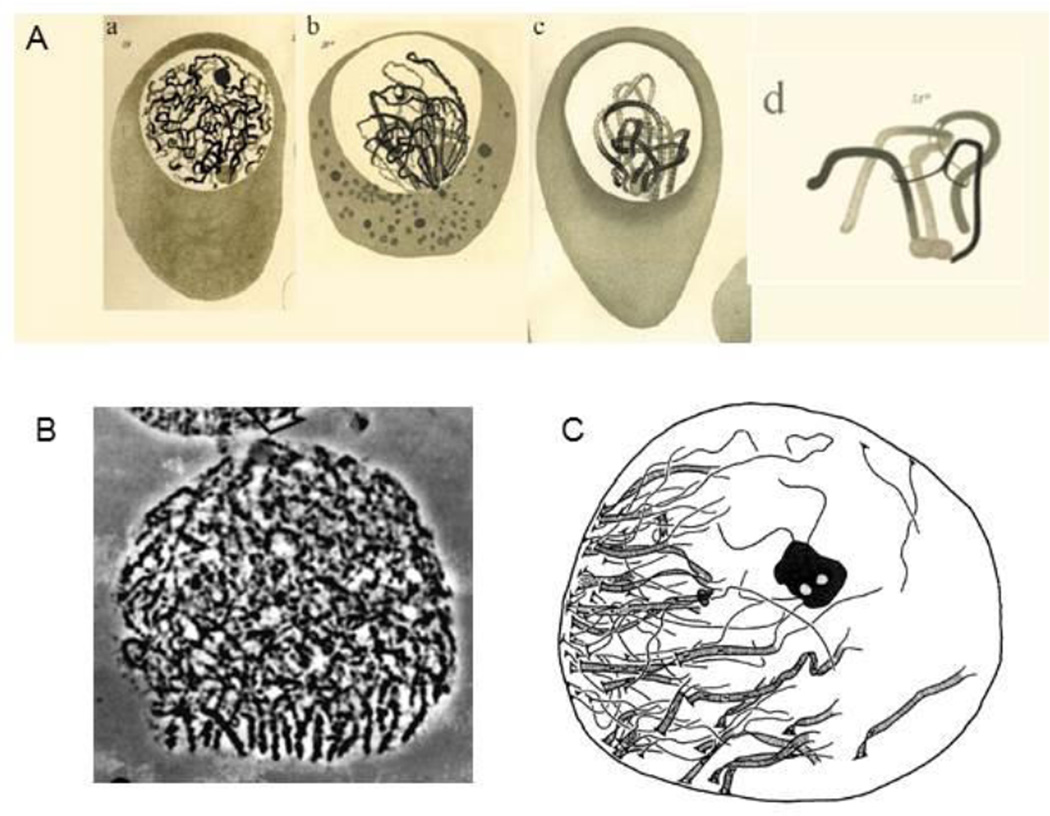
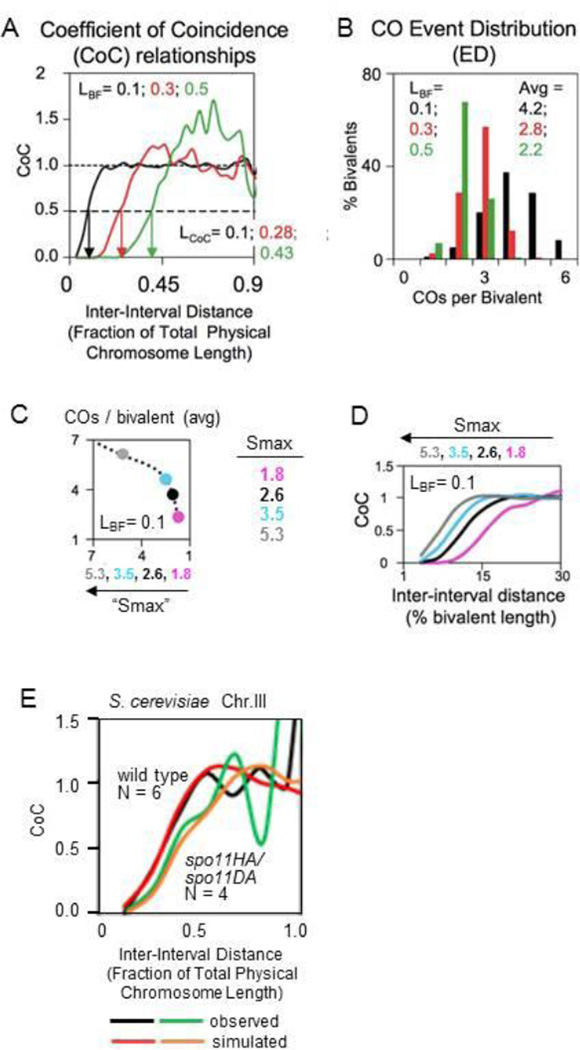
Meiosis presents many important mysteries that await elucidation. Here we discuss two such aspects. First, we consider how the current meiotic program might have evolved. We emphasize the central feature of this program: how homologous chromosomes find one another ("pair") so as to create the connections required for their regular segregation at Meiosis I. Points of emphasis include the facts that: (i) the classical "bouquet stage" is not required for initial homolog contacts in the current evolved meiotic program; and (ii) diverse observations point to commonality between molecules that mediate meiotic inter-homolog interactions and molecules that are integral to centromeres and/or to microtubule organizing centers (a.k.a. spindle pole bodies or centrosomes). Second, we provide an overview of the classical phenomenon of crossover (CO) interference in an effort to bridge the gap between description on the one hand versus logic and mechanism on the other.
Keywords: Bouquet; Crossover interference; Meiosis; Pairing.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
