

Circadian rhythms in neuronal activity propagate through output circuits
- PMID: 26928065
- PMCID: PMC5066395
- DOI: 10.1038/nn.4263
Circadian rhythms in neuronal activity propagate through output circuits
Abstract
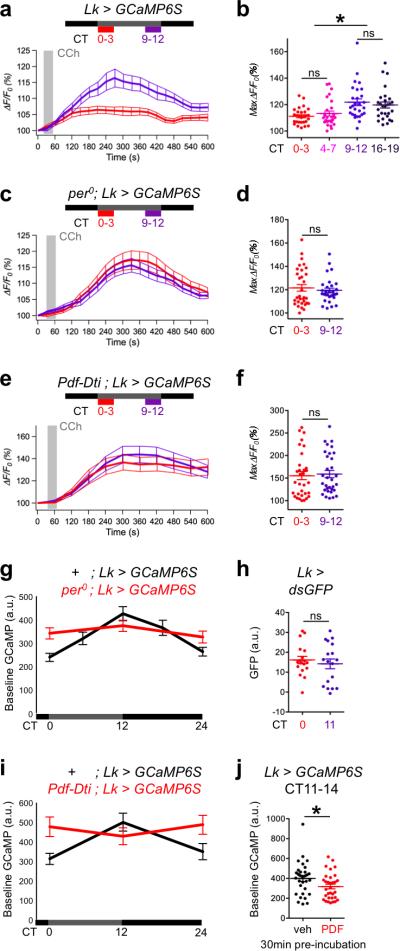
Twenty-four hour rhythms in behavior are organized by a network of circadian pacemaker neurons. Rhythmic activity in this network is generated by intrinsic rhythms in clock neuron physiology and communication between clock neurons. However, it is poorly understood how the activity of a small number of pacemaker neurons is translated into rhythmic behavior of the whole animal. To understand this, we screened for signals that could identify circadian output circuits in Drosophila melanogaster. We found that leucokinin neuropeptide (LK) and its receptor (LK-R) were required for normal behavioral rhythms. This LK/LK-R circuit connects pacemaker neurons to brain areas that regulate locomotor activity and sleep. Our experiments revealed that pacemaker neurons impose rhythmic activity and excitability on LK- and LK-R-expressing neurons. We also found pacemaker neuron-dependent activity rhythms in a second circadian output pathway controlled by DH44 neuropeptide-expressing neurons. We conclude that rhythmic clock neuron activity propagates to multiple downstream circuits to orchestrate behavioral rhythms.
Figures






Comment in
-
And the beat goes on: from clock to behavior.Nat Neurosci. 2016 Apr;19(4):529-30. doi: 10.1038/nn.4278. Nat Neurosci. 2016. PMID: 27021942 No abstract available.
References
-
- Herzog ED. Neurons and networks in daily rhythms. Nat Rev Neurosci. 2007;8:790–802. - PubMed
-
- Nitabach MN, Taghert PH. Organization of the Drosophila circadian control circuit. Curr Biol. 2008;18:R84–93. - PubMed
-
- Dibner C, Schibler U, Albrecht U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu Rev Physiol. 2010;72:517–549. - PubMed
Methods-only references
-
- Martinek S, Inonog S, Manoukian AS, Young MW. A role for the segment polarity gene shaggy/GSK-3 in the Drosophila circadian clock. Cell. 2001;105:769–779. - PubMed
-
- Plautz JD, Kaneko M, Hall JC, Kay SA. Independent photoreceptive circadian clocks throughout Drosophila. Science. 1997;278:1632–1635. - PubMed
-
- Calleja M, Moreno E, Pelaz S, Morata G. Visualization of gene expression in living adult Drosophila. Science. 1996;274:252–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
