

Interaction of extracellular S100A4 with RAGE prompts prometastatic activation of A375 melanoma cells
- PMID: 26928771
- PMCID: PMC4831350
- DOI: 10.1111/jcmm.12808
Interaction of extracellular S100A4 with RAGE prompts prometastatic activation of A375 melanoma cells
Abstract
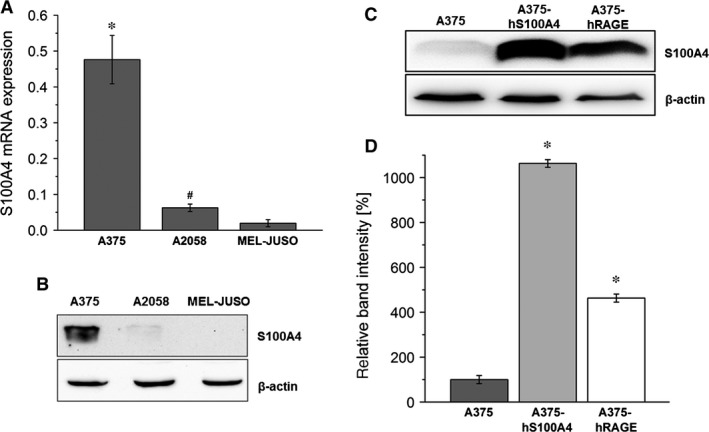
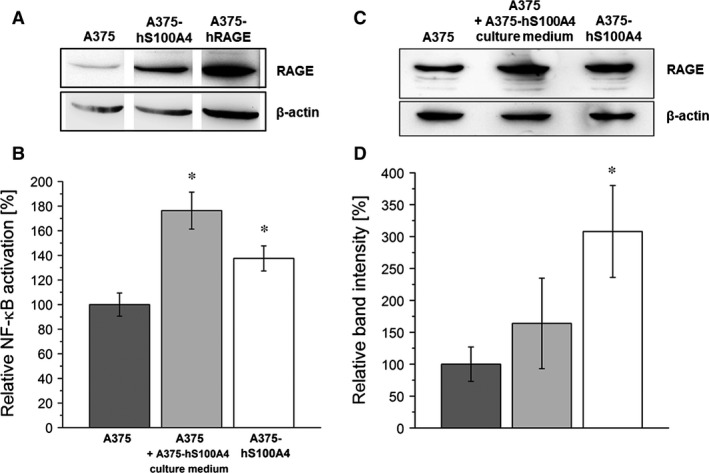
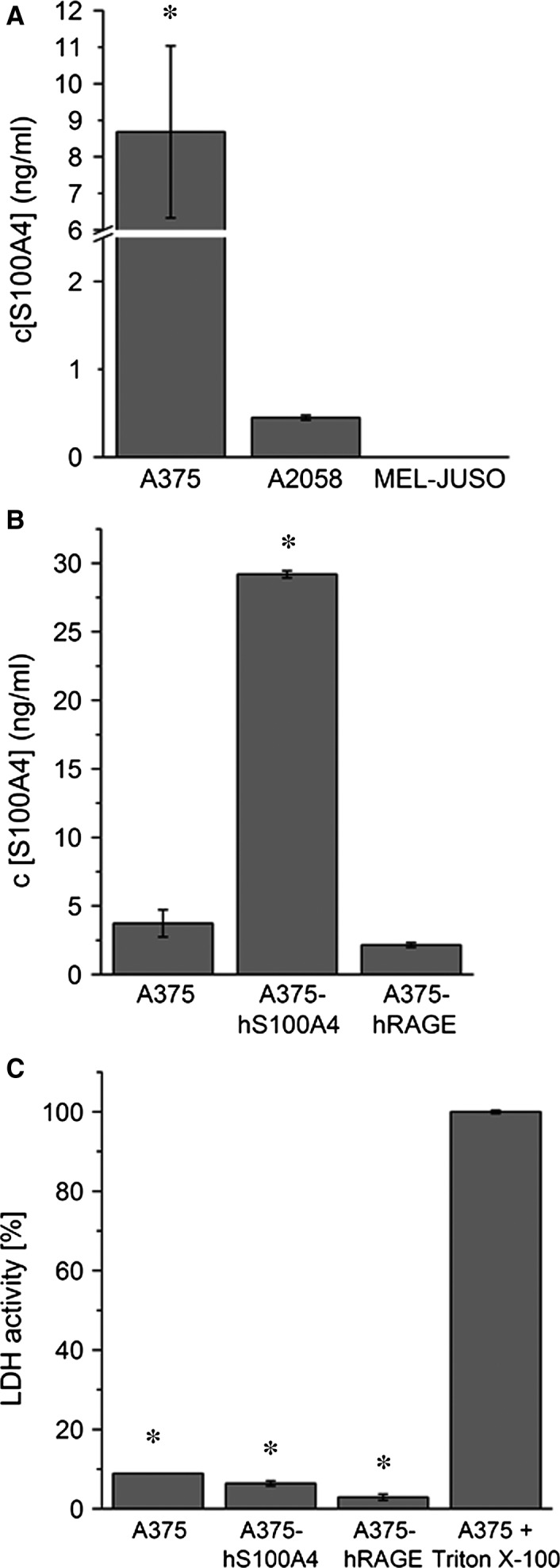
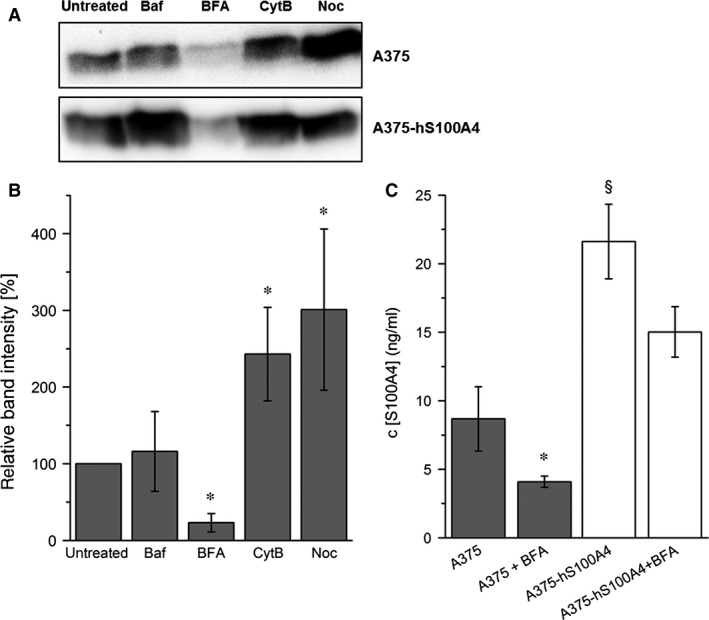
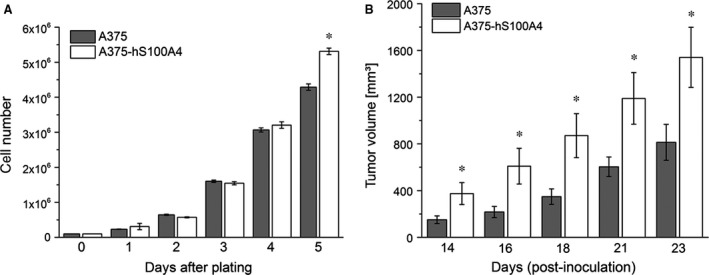
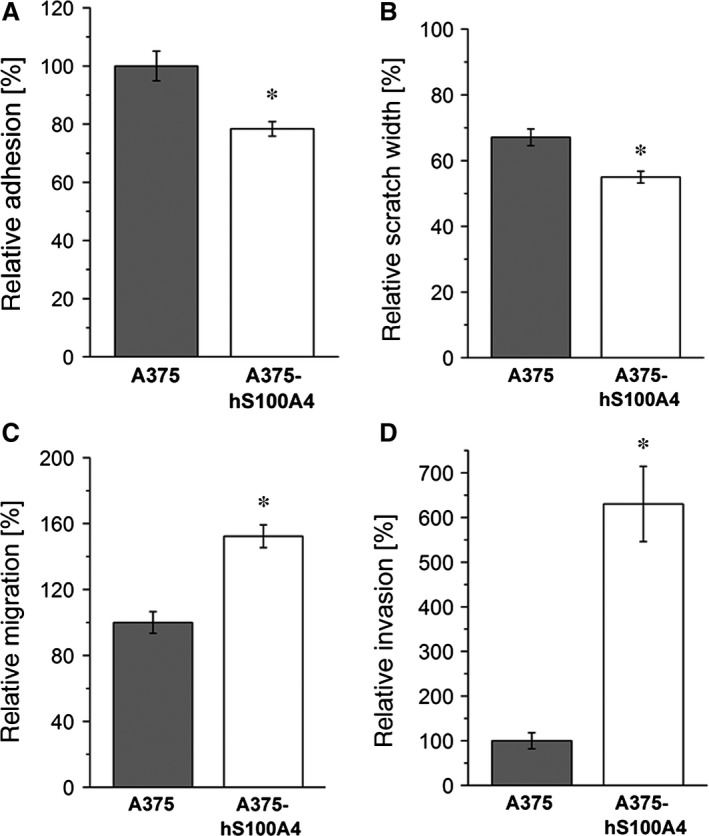
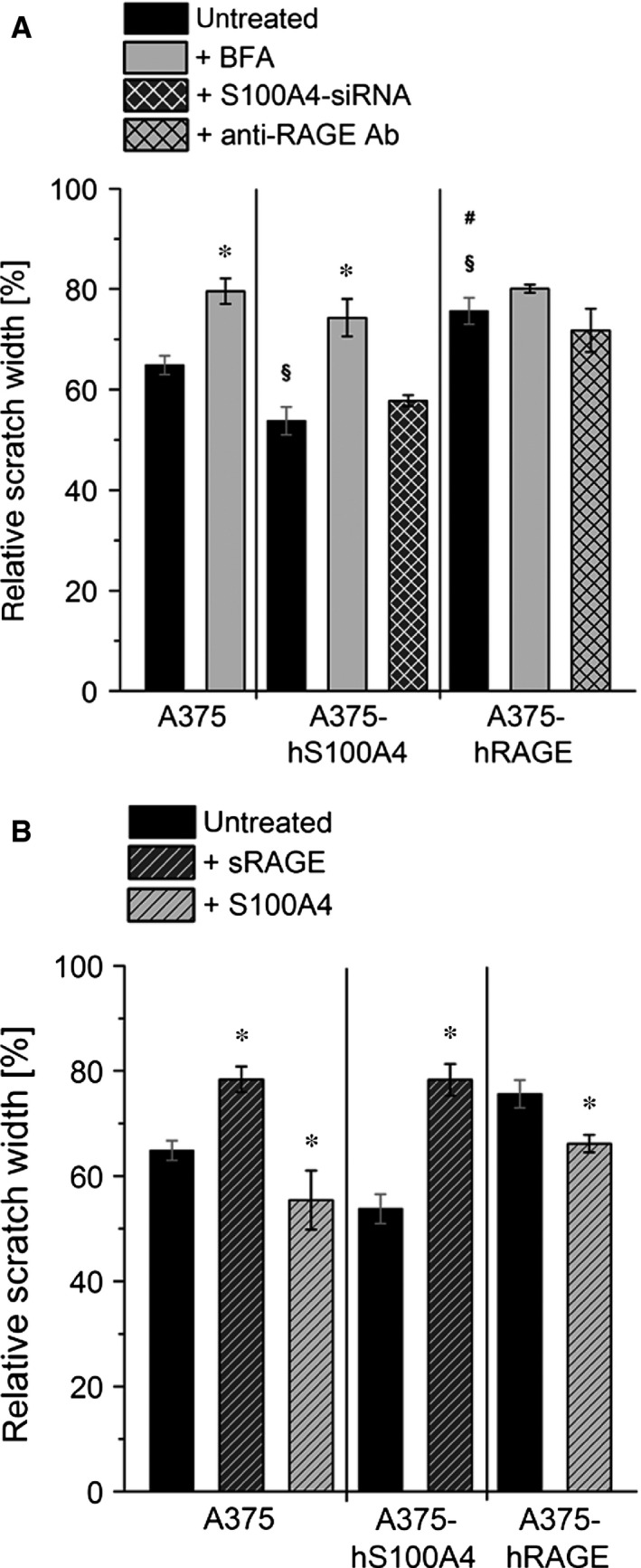
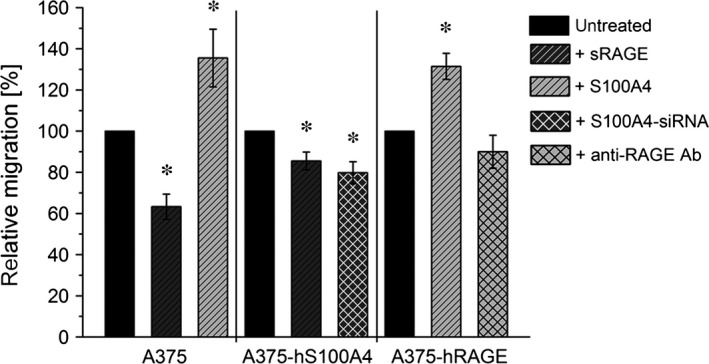
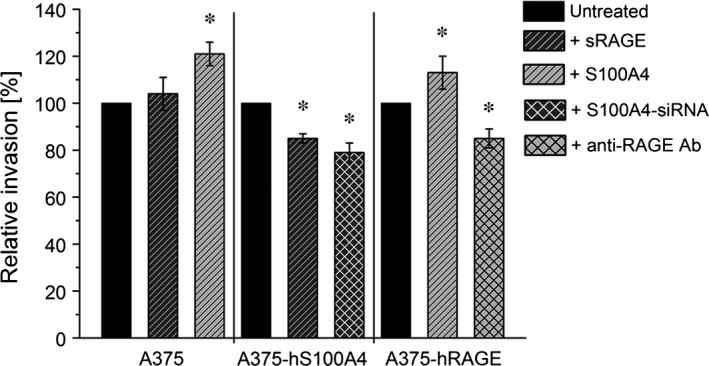
S100A4, a member of the S100 protein family of EF-hand calcium-binding proteins, is overexpressed in various tumour entities, including melanoma, and plays an important role in tumour progression. Several studies in epithelial and mesenchymal tumours revealed a correlation between extracellular S100A4 and metastasis. However, exact mechanisms how S100A4 stimulates metastasis in melanoma are still unknown. From a pilot experiment on baseline synthesis and secretion of S100A4 in human melanoma cell lines, which are in broad laboratory use, A375 wild-type cells and, additionally, newly generated A375 cell lines stably transfected with human S100A4 (A375-hS100A4) or human receptor for advanced glycation endproducts (A375-hRAGE), were selected to investigate the influence of extracellular S100A4 on cell motility, adhesion, migration and invasion in more detail. We demonstrated that A375 cells actively secrete S100A4 in the extracellular space via an endoplasmic reticulum-Golgi-dependent pathway. S100A4 overexpression and secretion resulted in prometastatic activation of A375 cells. Moreover, we determined the influence of S100A4-RAGE interaction and its blockade on A375, A375-hS100A4, A375-hRAGE cells, and showed that interaction of RAGE with extracellular S100A4 contributes to the observed activation of A375 cells. This investigation reveals additional molecular targets for therapeutic approaches aiming at blockade of ligand binding to RAGE or RAGE signalling to inhibit melanoma metastasis.
Keywords: ER-Golgi-dependent secretion pathway; S100 protein secretion; calcium-binding proteins; cancer metastasis; soluble receptor for advanced glycation endproducts.
© 2016 The Authors. Journal of Cellular and Molecular Medicine published by John Wiley & Sons Ltd and Foundation for Cellular and Molecular Medicine.
Figures
References
-
- Ko JM, Fisher DE. A new era: melanoma genetics and therapeutics. J Pathol. 2011; 223: 242–51. - PubMed
-
- Tandler N, Mosch B, Pietzsch J. Protein and non‐protein biomarkers in melanoma: a critical update. Amino Acids. 2012; 43: 2203–30. - PubMed
-
- Garrett SC, Varney KM, Weber DJ, et al S100A4, a mediator of metastasis. J Biol Chem. 2006; 281: 677–80. - PubMed
-
- Wolf S, Haase‐Kohn C, Lenk J, et al Expression, purification and fluorine‐18 radiolabeling of recombinant S100A4: a potential probe for molecular imaging of receptor for advanced glycation endproducts in vivo? Amino Acids. 2011; 41: 809–20. - PubMed
-
- Maelandsmo GM, Flørenes VA, Mellingsaeter T, et al Differential expression patterns of S100A2, S100A4 and S100A6 during progression of human malignant melanoma. Int J Cancer. 1997; 74: 464–9. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
