

Novel genomic island modifies DNA with 7-deazaguanine derivatives
- PMID: 26929322
- PMCID: PMC4801273
- DOI: 10.1073/pnas.1518570113
Novel genomic island modifies DNA with 7-deazaguanine derivatives
Abstract
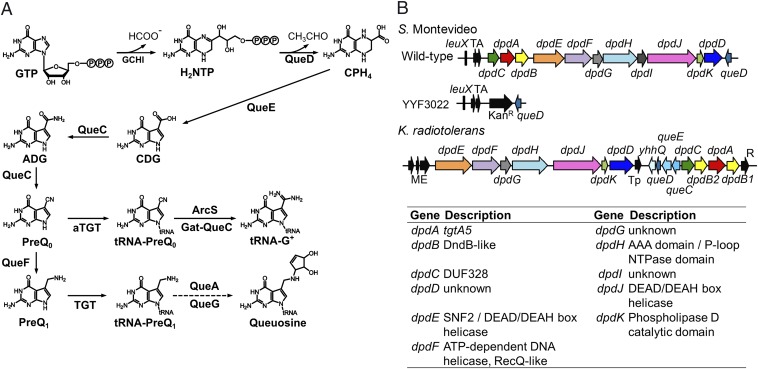
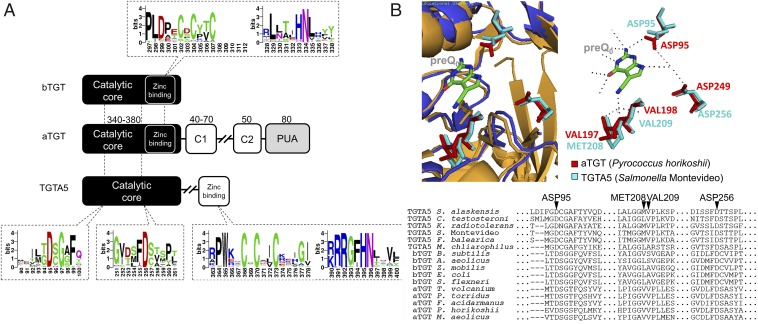
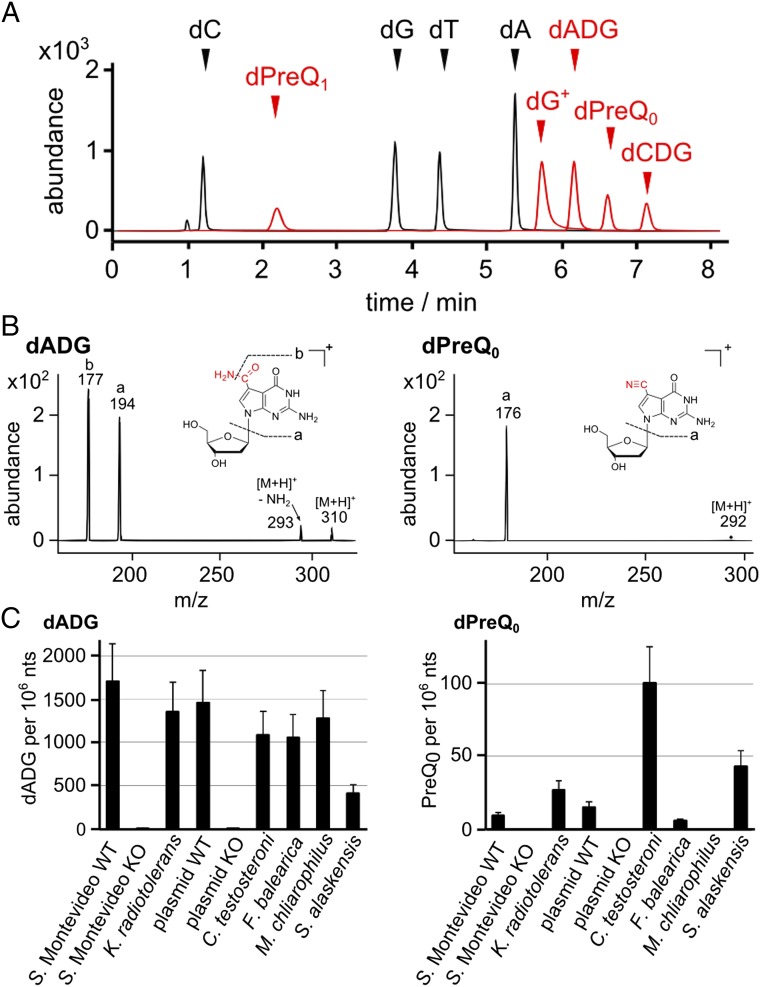
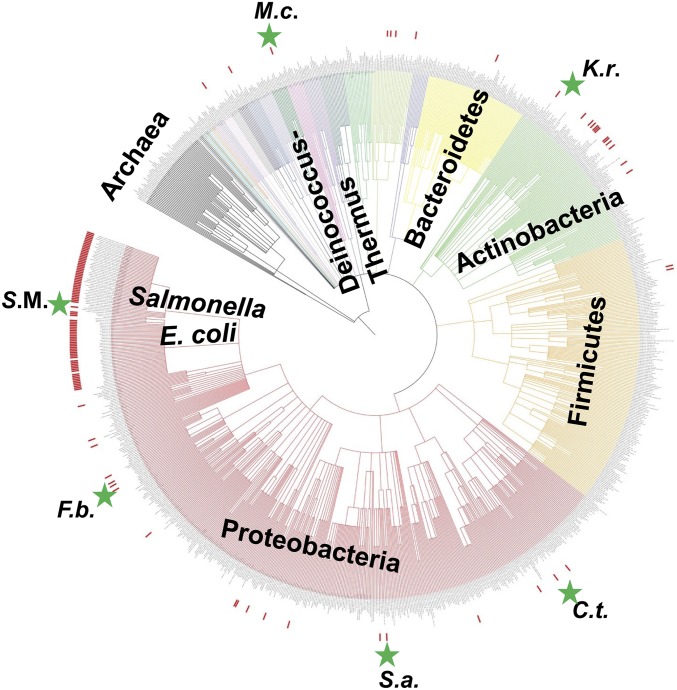
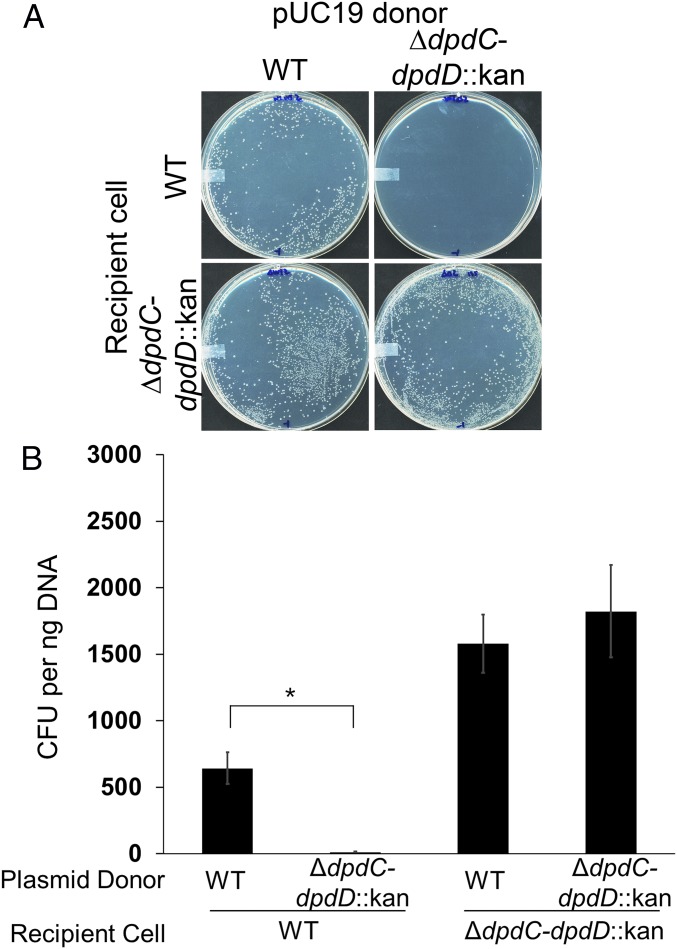
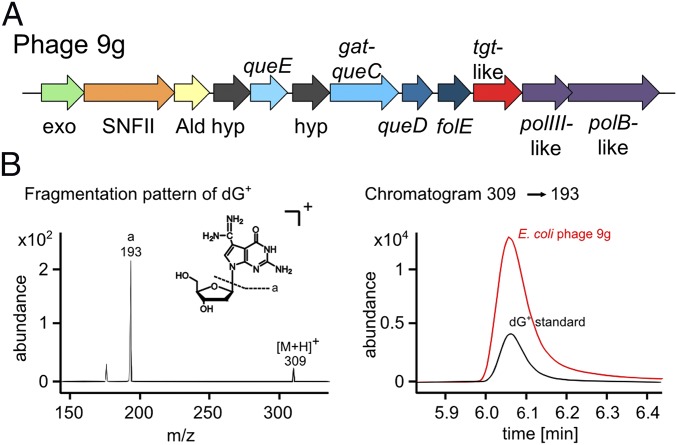
The discovery of ∼20-kb gene clusters containing a family of paralogs of tRNA guanosine transglycosylase genes, called tgtA5, alongside 7-cyano-7-deazaguanine (preQ0) synthesis and DNA metabolism genes, led to the hypothesis that 7-deazaguanine derivatives are inserted in DNA. This was established by detecting 2'-deoxy-preQ0 and 2'-deoxy-7-amido-7-deazaguanosine in enzymatic hydrolysates of DNA extracted from the pathogenic, Gram-negative bacteria Salmonella enterica serovar Montevideo. These modifications were absent in the closely related S. enterica serovar Typhimurium LT2 and from a mutant of S Montevideo, each lacking the gene cluster. This led us to rename the genes of the S. Montevideo cluster as dpdA-K for 7-deazapurine in DNA. Similar gene clusters were analyzed in ∼150 phylogenetically diverse bacteria, and the modifications were detected in DNA from other organisms containing these clusters, including Kineococcus radiotolerans, Comamonas testosteroni, and Sphingopyxis alaskensis Comparative genomic analysis shows that, in Enterobacteriaceae, the cluster is a genomic island integrated at the leuX locus, and the phylogenetic analysis of the TgtA5 family is consistent with widespread horizontal gene transfer. Comparison of transformation efficiencies of modified or unmodified plasmids into isogenic S. Montevideo strains containing or lacking the cluster strongly suggests a restriction-modification role for the cluster in Enterobacteriaceae. Another preQ0 derivative, 2'-deoxy-7-formamidino-7-deazaguanosine, was found in the Escherichia coli bacteriophage 9 g, as predicted from the presence of homologs of genes involved in the synthesis of the archaeosine tRNA modification. These results illustrate a deep and unexpected evolutionary connection between DNA and tRNA metabolism.
Keywords: 7-deazaguanine; DNA modification; comparative genomics; queuosine; restriction–modification.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Grosjean H. Nucleic acids are not boring long polymers of only four types of nucleotides. In: Grosjean H, editor. DNA and RNA Modification Enzymes: Structure, Mechanism, Function and Evolution. Landes Bioscience; Austin, TX: 2009. pp. 1–18.
-
- Warren RA. Modified bases in bacteriophage DNAs. Annu Rev Microbiol. 1980;34:137–158. - PubMed
-
- Wang L, et al. Phosphorothioation of DNA in bacteria by dnd genes. Nat Chem Biol. 2007;3(11):709–710. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
