

Functional and topological diversity of LOV domain photoreceptors
- PMID: 26929367
- PMCID: PMC4801262
- DOI: 10.1073/pnas.1509428113
Functional and topological diversity of LOV domain photoreceptors
Abstract
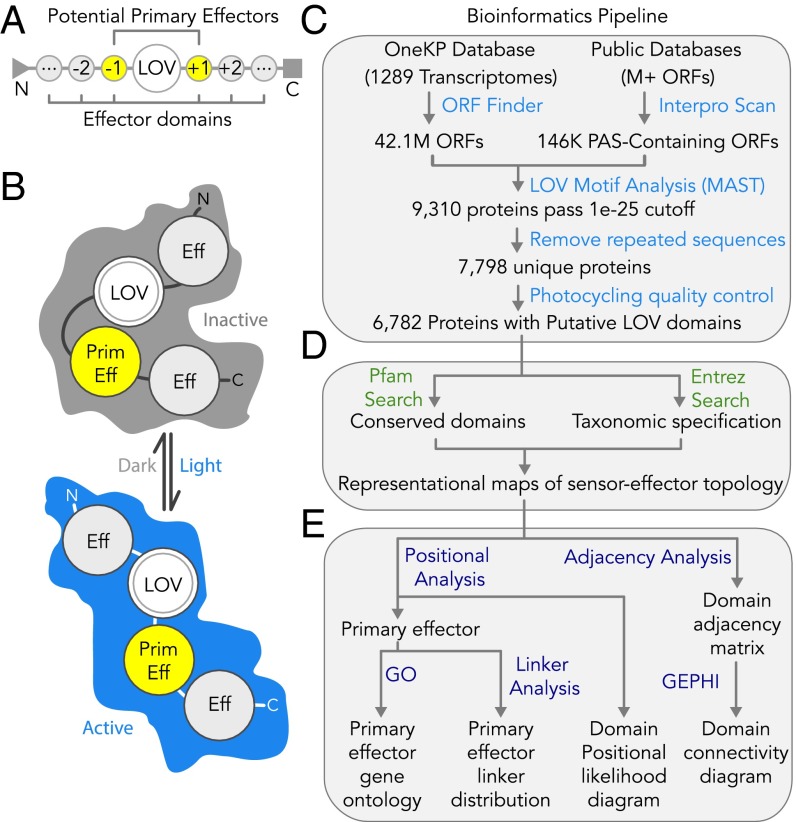
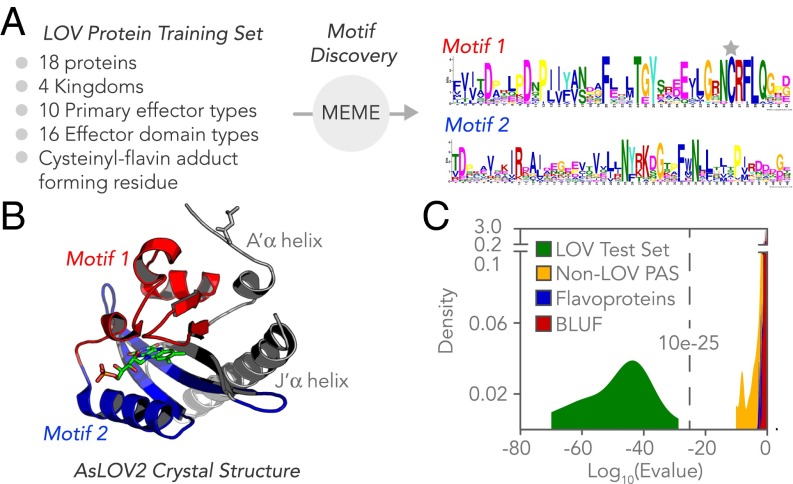
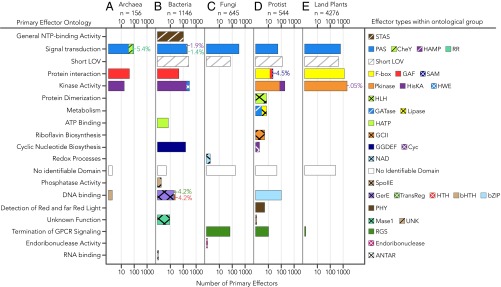
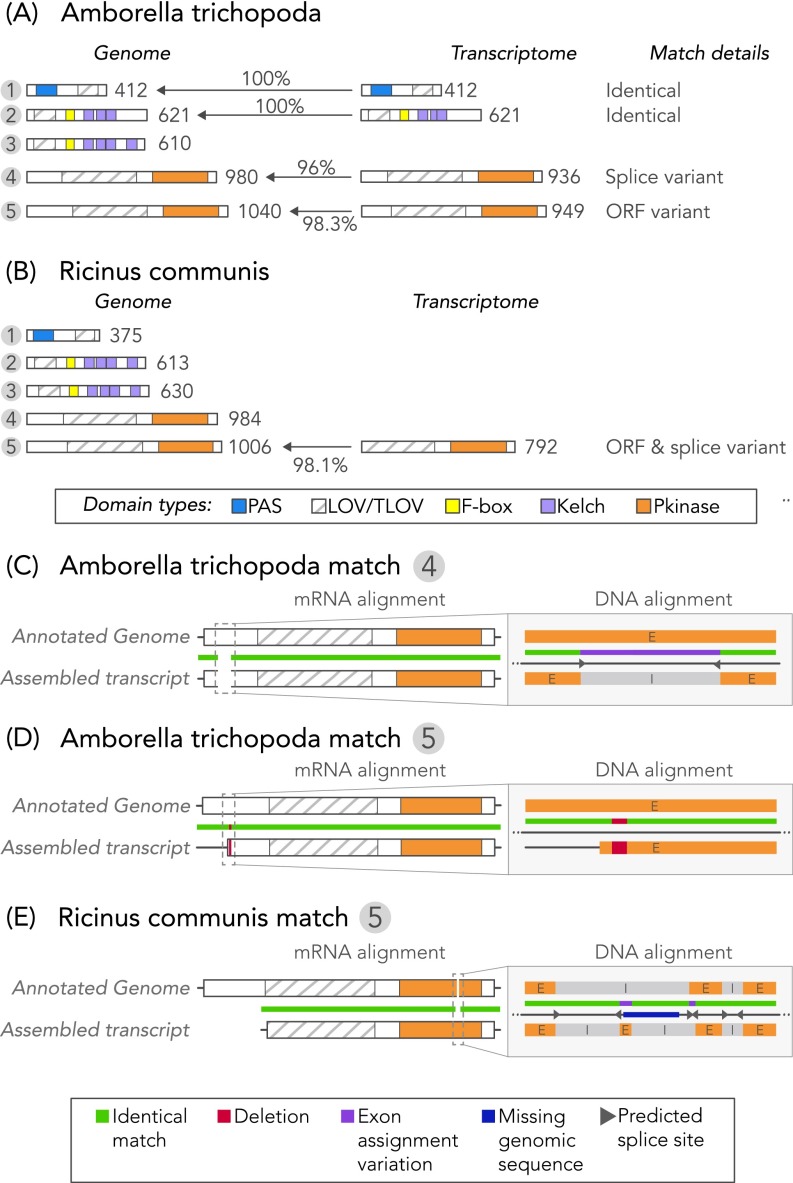
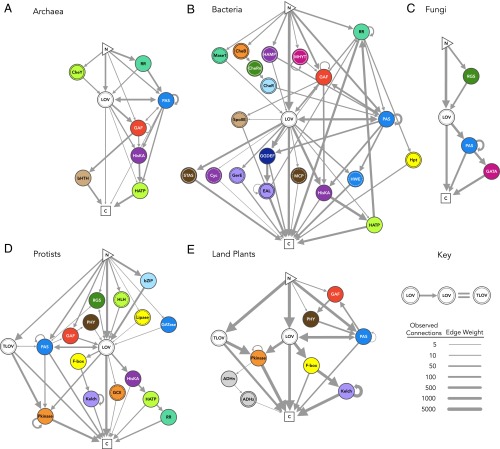
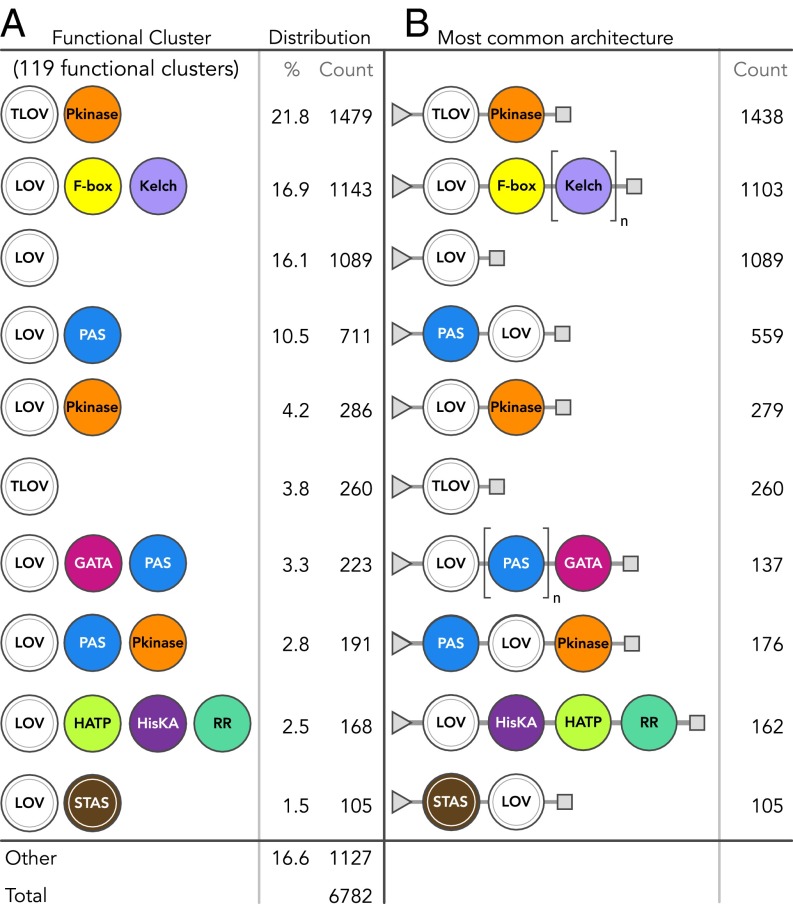
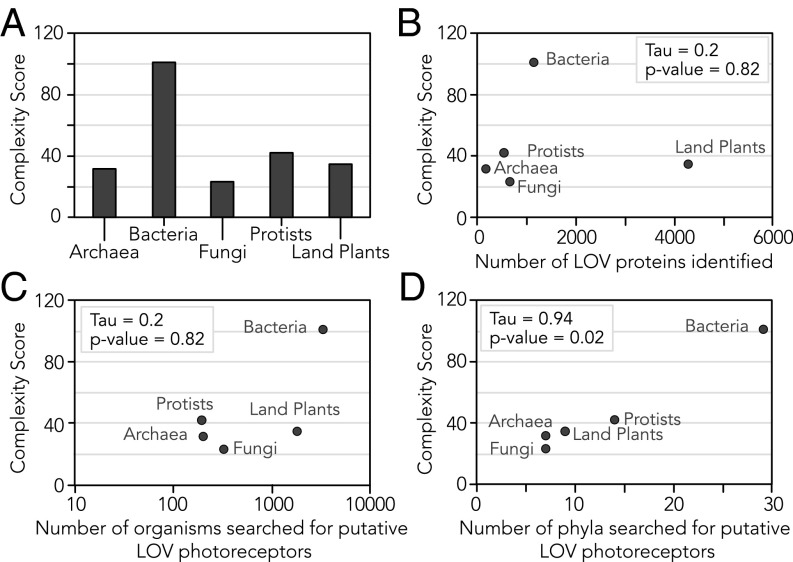
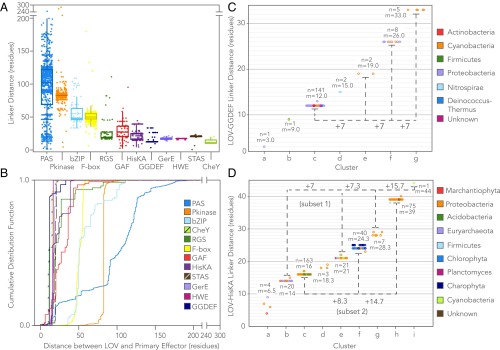
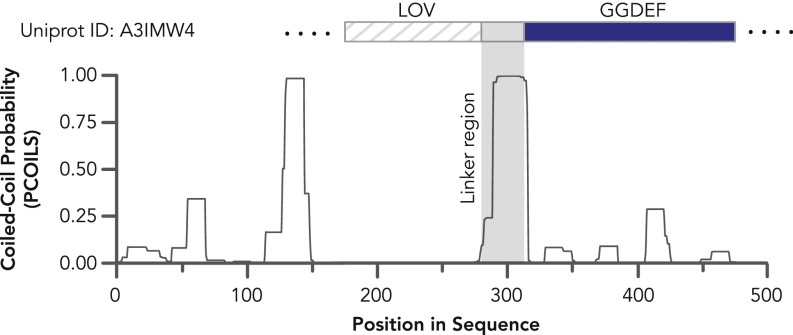
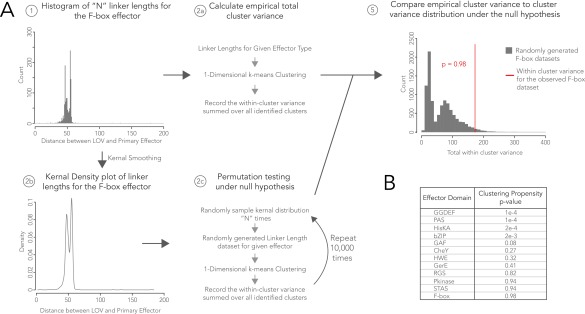
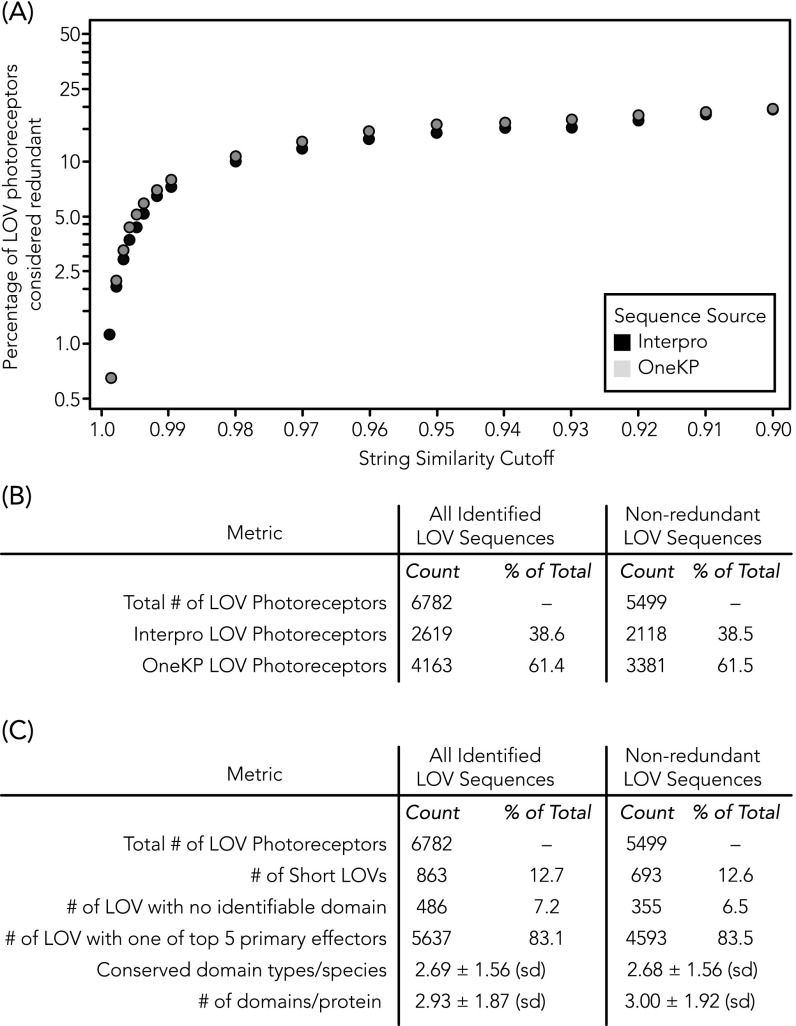
Light-oxygen-voltage sensitive (LOV) flavoproteins are ubiquitous photoreceptors that mediate responses to environmental cues. Photosensory inputs are transduced into signaling outputs via structural rearrangements in sensor domains that consequently modulate the activity of an effector domain or multidomain clusters. Establishing the diversity in effector function and sensor-effector topology will inform what signaling mechanisms govern light-responsive behaviors across multiple kingdoms of life and how these signals are transduced. Here, we report the bioinformatics identification of over 6,700 candidate LOV domains (including over 4,000 previously unidentified sequences from plants and protists), and insights from their annotations for ontological function and structural arrangements. Motif analysis identified the sensors from ∼42 million ORFs, with strong statistical separation from other flavoproteins and non-LOV members of the structurally related Per-aryl hydrocarbon receptor nuclear translocator (ARNT)-Sim family. Conserved-domain analysis determined putative light-regulated function and multidomain topologies. We found that for certain effectors, sensor-effector linker length is discretized based on both phylogeny and the preservation of α-helical heptad repeats within an extended coiled-coil linker structure. This finding suggests that preserving sensor-effector orientation is a key determinant of linker length, in addition to ancestry, in LOV signaling structure-function. We found a surprisingly high prevalence of effectors with functions previously thought to be rare among LOV proteins, such as regulators of G protein signaling, and discovered several previously unidentified effectors, such as lipases. This work highlights the value of applying genomic and transcriptomic technologies to diverse organisms to capture the structural and functional variation in photosensory proteins that are vastly important in adaptation, photobiology, and optogenetics.
Keywords: LOV; flavoproteins; optogenetics; photoreceptors.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Crosson S, Rajagopal S, Moffat K. The LOV domain family: Photoresponsive signaling modules coupled to diverse output domains. Biochemistry. 2003;42(1):2–10. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R21 DA040434/DA/NIDA NIH HHS/United States
- R01 NS075421/NS/NINDS NIH HHS/United States
- R01 NS067199/NS/NINDS NIH HHS/United States
- R01 DA029639/DA/NIDA NIH HHS/United States
- RC1 MH088182/MH/NIMH NIH HHS/United States
- 1R21 DA040434-01/DA/NIDA NIH HHS/United States
- 1R01NS075421/NS/NINDS NIH HHS/United States
- DP2 OD002002/OD/NIH HHS/United States
- R01 GM104948/GM/NIGMS NIH HHS/United States
- R01 GM106239/GM/NIGMS NIH HHS/United States
- 1RC1MH088182/MH/NIMH NIH HHS/United States
- 1R01DA029639/DA/NIDA NIH HHS/United States
- 1R01GM104948/GM/NIGMS NIH HHS/United States
- 1DP2OD002002/OD/NIH HHS/United States
- 1R01NS067199/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
