

Adhesion protein networks reveal functions proximal and distal to cell-matrix contacts
- PMID: 26930633
- PMCID: PMC5094910
- DOI: 10.1016/j.ceb.2016.02.013
Adhesion protein networks reveal functions proximal and distal to cell-matrix contacts
Abstract
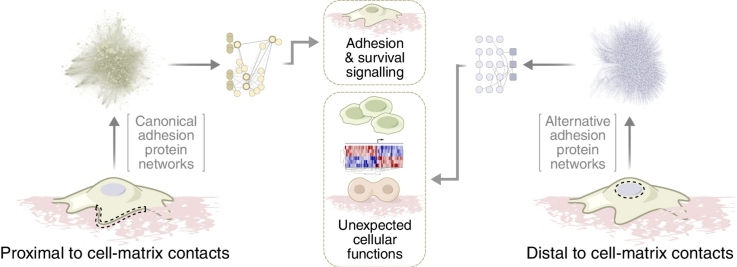
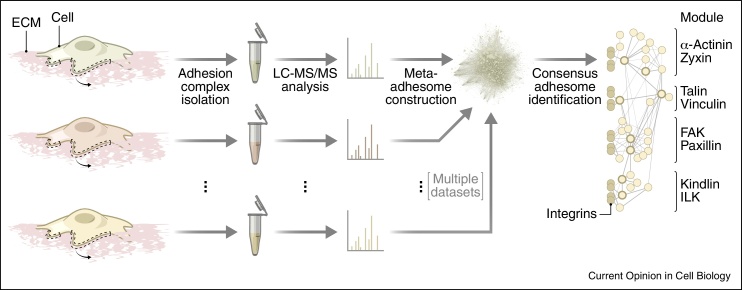
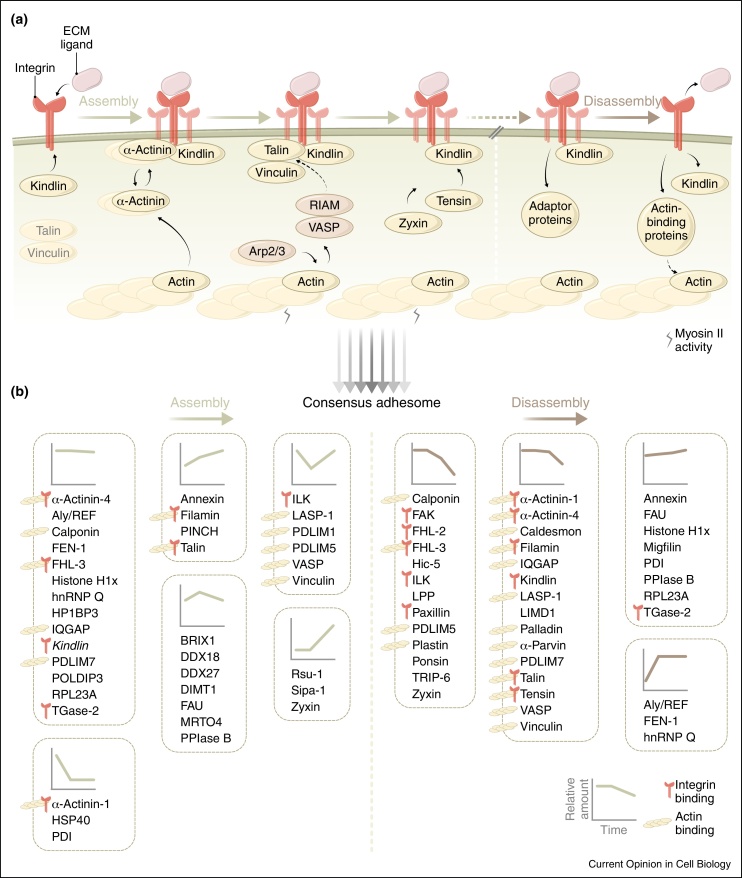
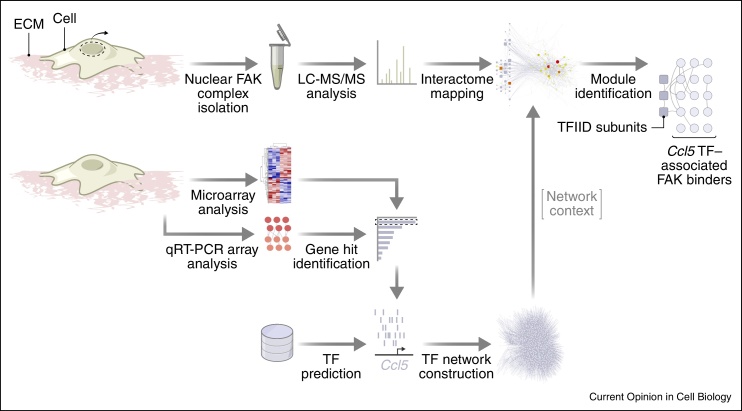
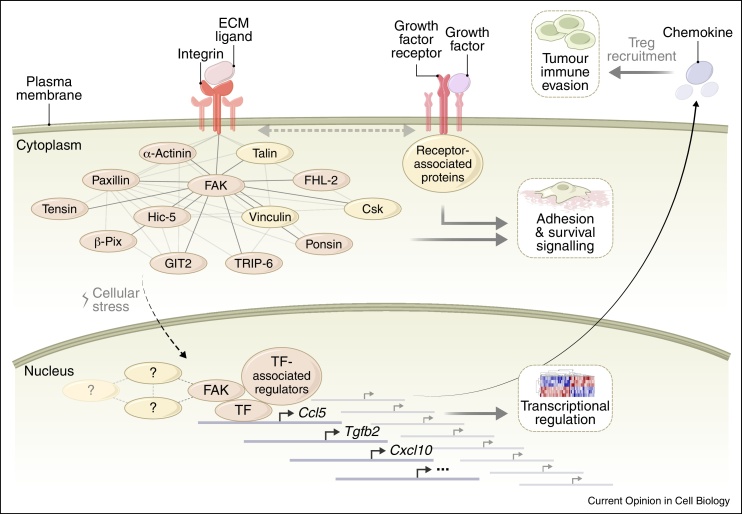
Cell adhesion to the extracellular matrix is generally mediated by integrin receptors, which bind to intracellular adhesion proteins that form multi-molecular scaffolding and signalling complexes. The networks of proteins, and their interactions, are dynamic, mechanosensitive and extremely complex. Recent efforts to characterise adhesions using a variety of technologies, including imaging, proteomics and bioinformatics, have provided new insights into their composition, organisation and how they are regulated, and have also begun to reveal unexpected roles for so-called adhesion proteins in other cellular compartments (for example, the nucleus or centrosomes) in diseases such as cancer. We believe this is opening a new chapter on understanding the wider functions of adhesion proteins, both proximal and distal to cell-matrix contacts.
Copyright © 2016 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Charras G., Sahai E. Physical influences of the extracellular environment on cell migration. Nat Rev Mol Cell Biol. 2014;15:813–824. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
