

AMP-activated protein kinase: a cellular energy sensor that comes in 12 flavours
- PMID: 26934201
- PMCID: PMC4995730
- DOI: 10.1111/febs.13698
AMP-activated protein kinase: a cellular energy sensor that comes in 12 flavours
Abstract
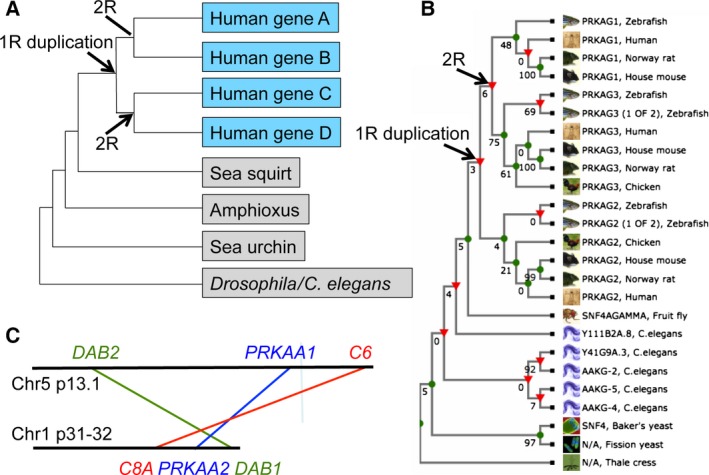
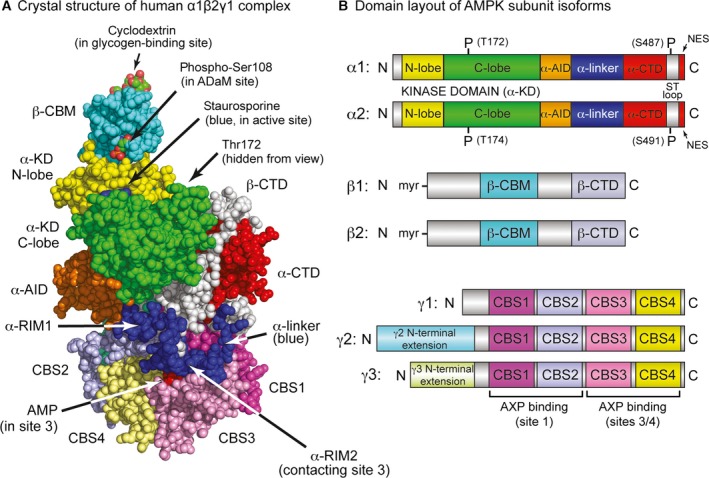
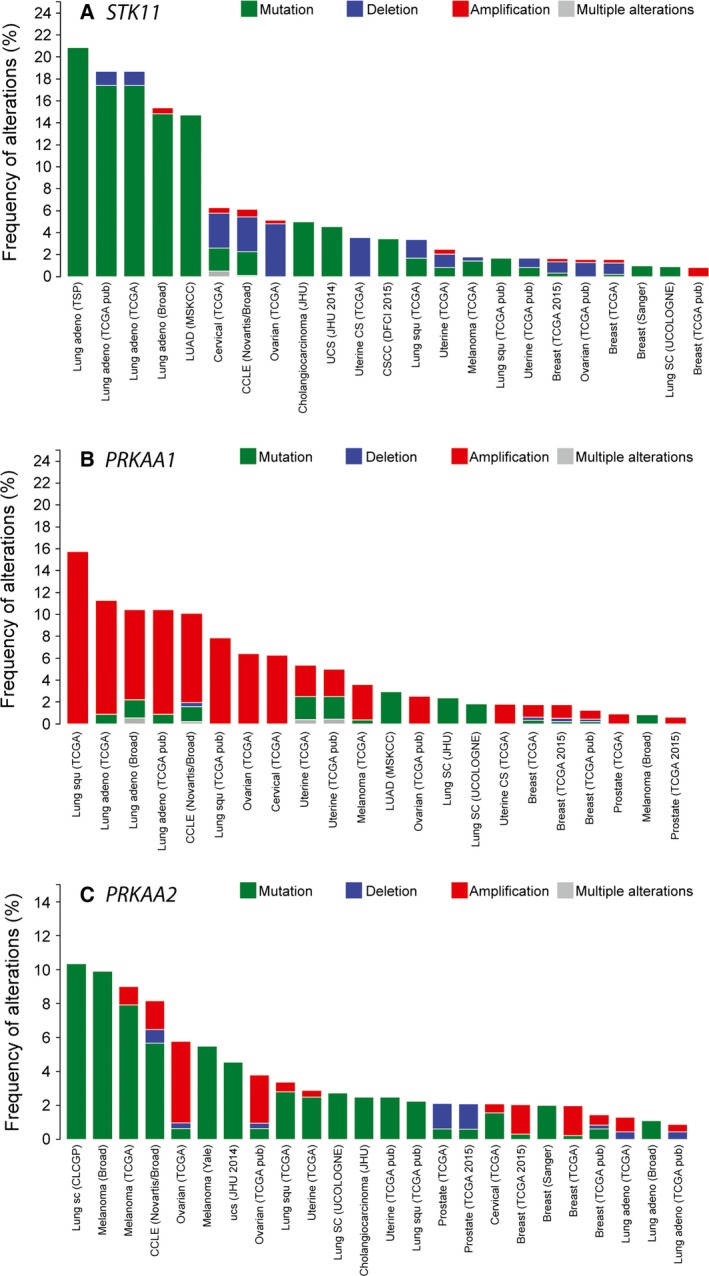
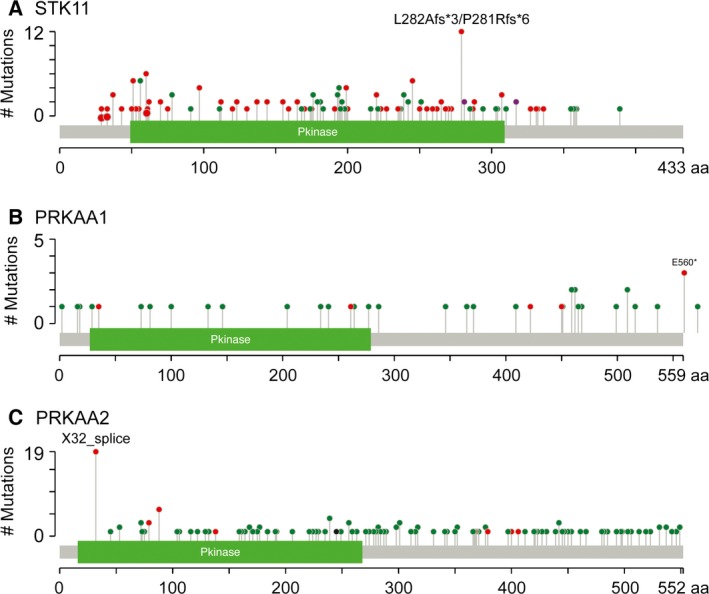
The AMP-activated protein kinase (AMPK) is a sensor of cellular energy status that is expressed in essentially all eukaryotic cells, suggesting that it arose during early eukaryotic evolution. It occurs universally as heterotrimeric complexes containing catalytic α subunits and regulatory β and γ subunits. Although Drosophila melanogaster contains single genes encoding each subunit, in mammals, each subunit exists as multiple isoforms encoded by distinct genes, giving rise to up to 12 heterotrimeric combinations. The multiple isoforms of each subunit are 2R-ohnologues generated by the two rounds of whole genome duplication that occurred at the evolutionary origin of the vertebrates. Although the differential roles of these isoform combinations remain only partly understood, there are indications that they may have different subcellular locations, different inputs and outputs, and different functions. The multiple isoforms are of particular interest with respect to the roles of AMPK in cancer because the genes encoding some isoforms, such as PRKAA1 and PRKAB2 (encoding α1 and β2), are quite frequently amplified in tumour cells, whereas the genes encoding others, such as PRKAA2 (encoding α2), tend to be mutated, which, in some but not all cases, may result in a loss of function. Thus, although AMPK acts downstream of the tumour suppressor liver kinase B1, and some of its isoform combinations may act as tumour suppressors that restrain the growth and proliferation of tumour cells, other isoform combinations may paradoxically act as oncogenes, perhaps by aiding the survival of tumour cells undergoing environmental stresses such as hypoxia or nutrient deprivation.
Keywords: 2R-ohnologue; AMP-activated protein kinase; LKB1; adenine nucleotides; cancer; energy homeostasis; oncogene; tumour suppressor.
© 2016 The Authors. The FEBS Journal published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Figures
References
-
- Stapleton D, Mitchelhill KI, Gao G, Widmer J, Michell BJ, Teh T, House CM, Fernandez CS, Cox T, Witters LA et al (1996) Mammalian AMP‐activated protein kinase subfamily. J Biol Chem 271, 611–614. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
