

Myosin filament sliding through the Z-disc relates striated muscle fibre structure to function
- PMID: 26936248
- PMCID: PMC4810864
- DOI: 10.1098/rspb.2015.3030
Myosin filament sliding through the Z-disc relates striated muscle fibre structure to function
Abstract
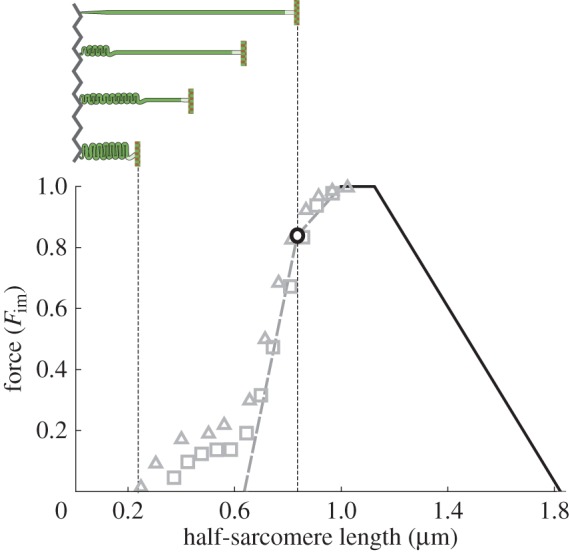
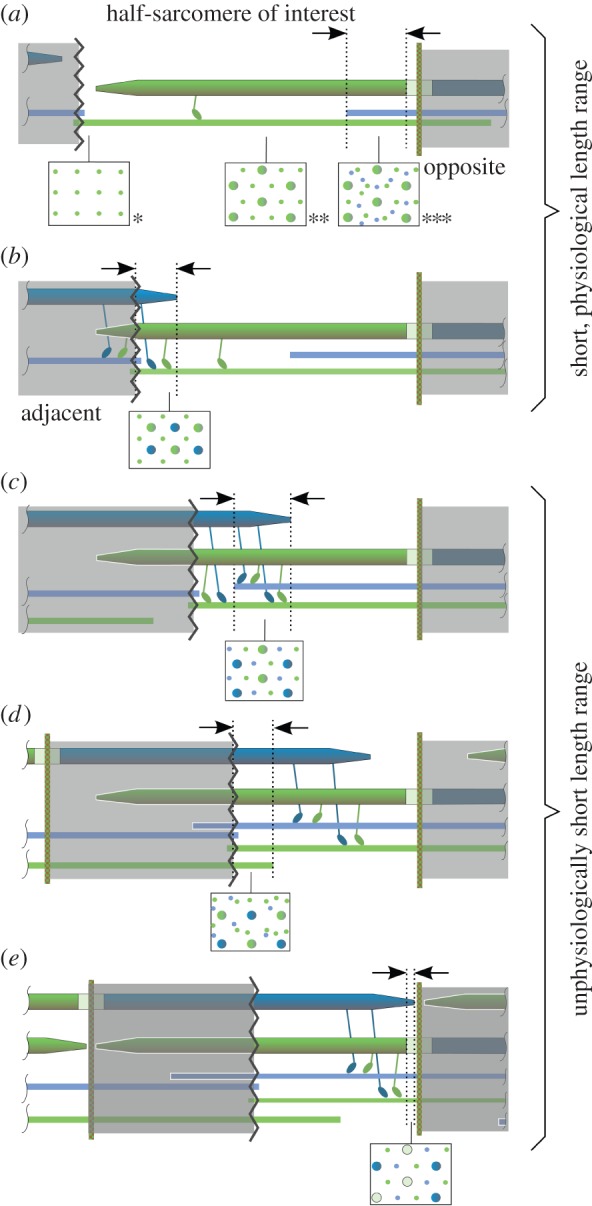
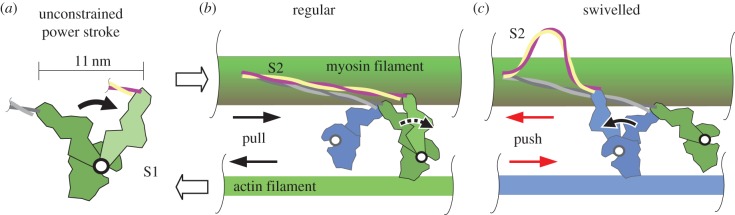
Striated muscle contraction requires intricate interactions of microstructures. The classic textbook assumption that myosin filaments are compressed at the meshed Z-disc during striated muscle fibre contraction conflicts with experimental evidence. For example, myosin filaments are too stiff to be compressed sufficiently by the muscular force, and, unlike compressed springs, the muscle fibres do not restore their resting length after contractions to short lengths. Further, the dependence of a fibre's maximum contraction velocity on sarcomere length is unexplained to date. In this paper, we present a structurally consistent model of sarcomere contraction that reconciles these findings with the well-accepted sliding filament and crossbridge theories. The few required model parameters are taken from the literature or obtained from reasoning based on structural arguments. In our model, the transition from hexagonal to tetragonal actin filament arrangement near the Z-disc together with a thoughtful titin arrangement enables myosin filament sliding through the Z-disc. This sliding leads to swivelled crossbridges in the adjacent half-sarcomere that dampen contraction. With no fitting of parameters required, the model predicts straightforwardly the fibre's entire force-length behaviour and the dependence of the maximum contraction velocity on sarcomere length. Our model enables a structurally and functionally consistent view of the contractile machinery of the striated fibre with possible implications for muscle diseases and evolution.
Keywords: Z-disc; model; myofilaments; sliding filament theory; striated muscle fibre.
© 2016 The Author(s).
Figures




References
-
- Blix M. 1891. Die Länge und die Spannung des Muskels. Skand. Arch. Physiol. 3, 295–118. ( 10.1111/j.1748-1716.1892.tb00660.x) - DOI
-
- Ramsey RW, Street SF. 1940. The isometric length-tension diagram of isolated skeletal muscle fibers of the frog. J. Cell. Compar. Physiol. 15, 11–34. ( 10.1002/jcp.1030150103) - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources