

Interneurons Differentially Contribute to Spontaneous Network Activity in the Developing Hippocampus Dependent on Their Embryonic Lineage
- PMID: 26937006
- PMCID: PMC4879211
- DOI: 10.1523/JNEUROSCI.4000-15.2016
Interneurons Differentially Contribute to Spontaneous Network Activity in the Developing Hippocampus Dependent on Their Embryonic Lineage
Abstract
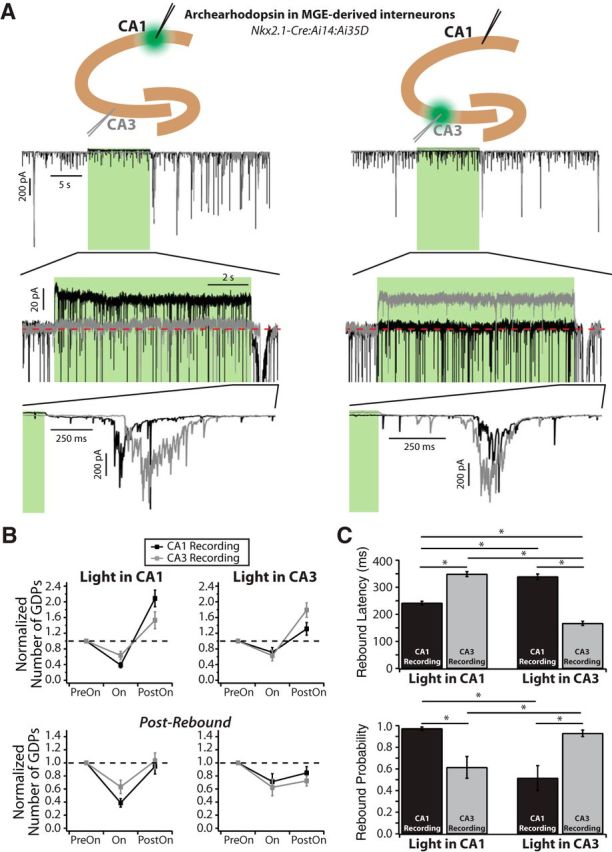
Spontaneously generated network activity is a hallmark of developing neural circuits, and plays an important role in the formation of synaptic connections. In the rodent hippocampus, this activity is observed in vitro as giant depolarizing potentials (GDPs) during the first postnatal week. Interneurons importantly contribute to GDPs, due to the depolarizing actions of GABA early in development. While they are highly diverse, cortical interneurons can be segregated into two distinct groups based on their embryonic lineage from either the medial or caudal ganglionic eminences (MGE and CGE). There is evidence suggesting CGE-derived interneurons are important for GDP generation; however, their contribution relative to those from the MGE has never been directly tested. Here, we optogenetically inhibited either MGE- or CGE-derived interneurons in a region-specific manner in mouse neonatal hippocampus in vitro. In CA1, where interneurons are the primary source of recurrent excitation, we found that those from the MGE strongly and preferentially contributed to GDP generation. Furthermore, in dual whole-cell patch recordings in neonatal CA1, MGE interneurons formed synaptic connections to and from neighboring pyramidal cells at a much higher rate than those from the CGE. These MGE interneurons were commonly perisomatic targeting, in contrast to those from the CGE, which were dendrite targeting. Finally, inhibiting MGE interneurons in CA1 suppressed GDPs in CA3 and vice versa; conversely, they could also trigger GDPs in CA1 that propagated to CA3 and vice versa. Our data demonstrate a key role for MGE-derived interneurons in both generating and coordinating GDPs across the hippocampus.
Significance statement: During nervous system development, immature circuits internally generate rhythmic patterns of electrical activity that promote the establishment of synaptic connections. Immature interneurons are excitatory rather than inhibitory and actively contribute to the generation of these spontaneous network events, referred to as giant depolarizing potentials (GDPs) in the hippocampus. Interneurons can be generally separated into two distinct groups based on their origin in the embryo from the medial or caudal ganglionic eminences (MGE and CGE). Here we show that MGE interneurons play a dominant role in generating GDPs compared with their CGE counterparts. They accomplish this due to their high synaptic connectivity within the local circuitry. Finally, they can control network activity across large regions of the developing hippocampus.
Keywords: giant depolarizing potential; hippocampus; interneurons.
Copyright © 2016 the authors 0270-6474/16/362646-17$15.00/0.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous