

Oral Immunization of Rabbits with S. enterica Typhimurium Expressing Neisseria gonorrhoeae Filamentous Phage Φ6 Induces Bactericidal Antibodies Against N. gonorrhoeae
- PMID: 26939573
- PMCID: PMC4778046
- DOI: 10.1038/srep22549
Oral Immunization of Rabbits with S. enterica Typhimurium Expressing Neisseria gonorrhoeae Filamentous Phage Φ6 Induces Bactericidal Antibodies Against N. gonorrhoeae
Abstract
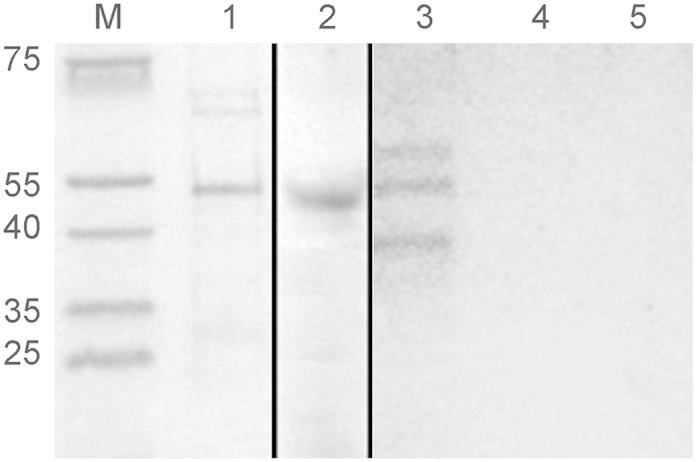
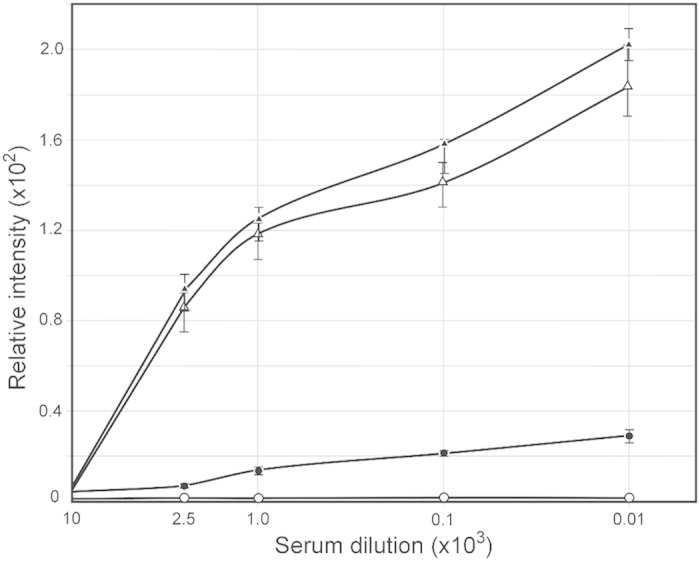
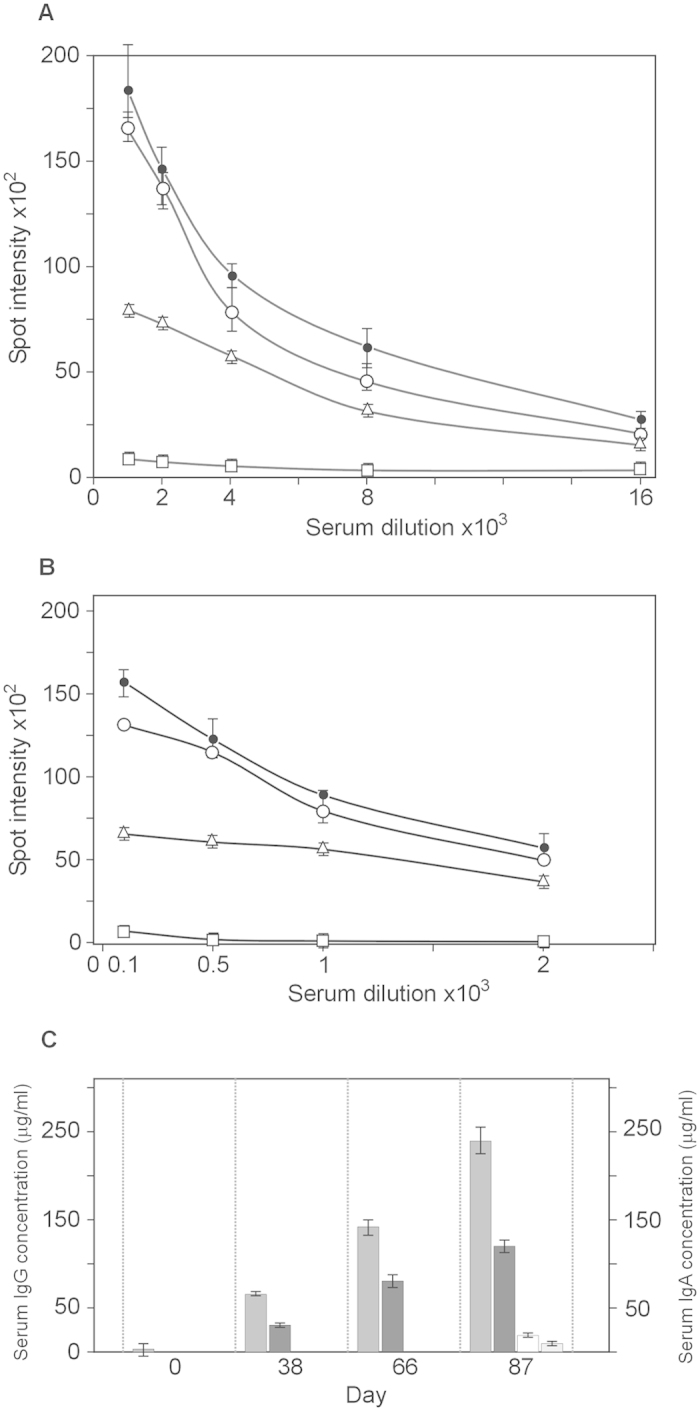
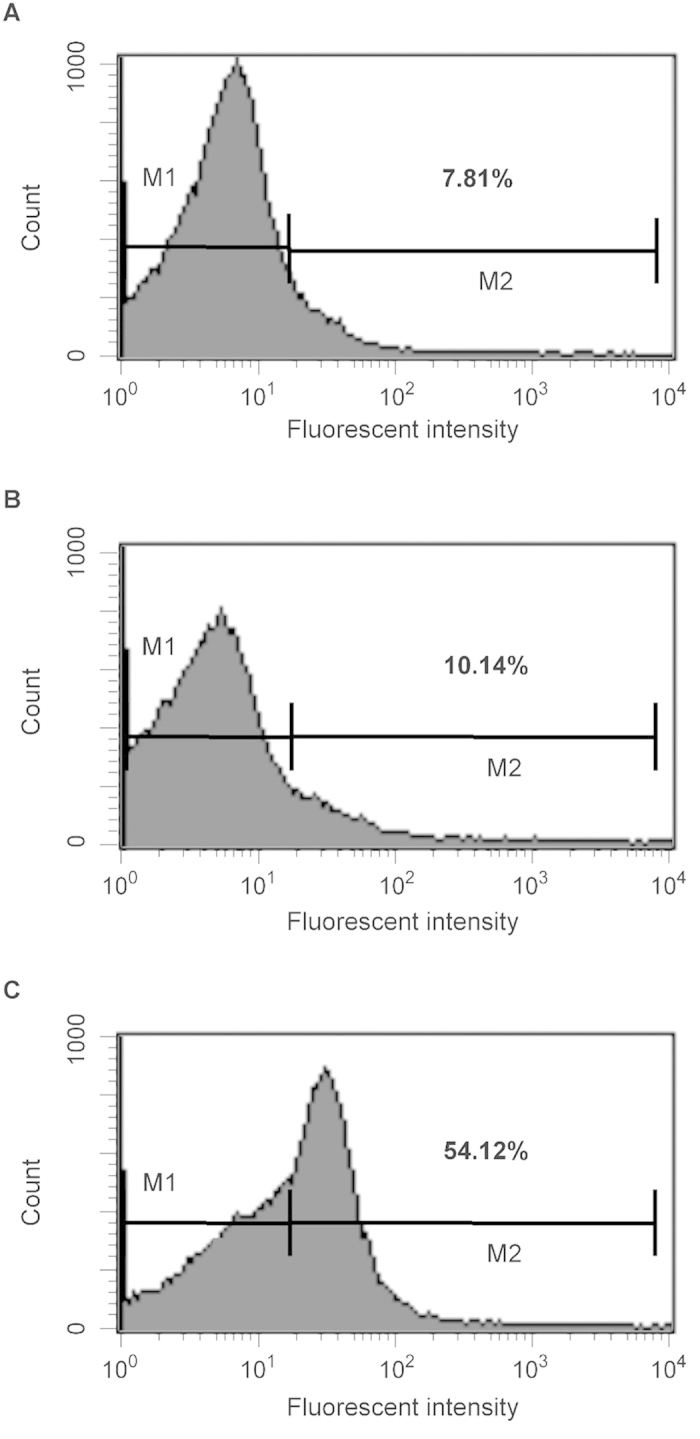
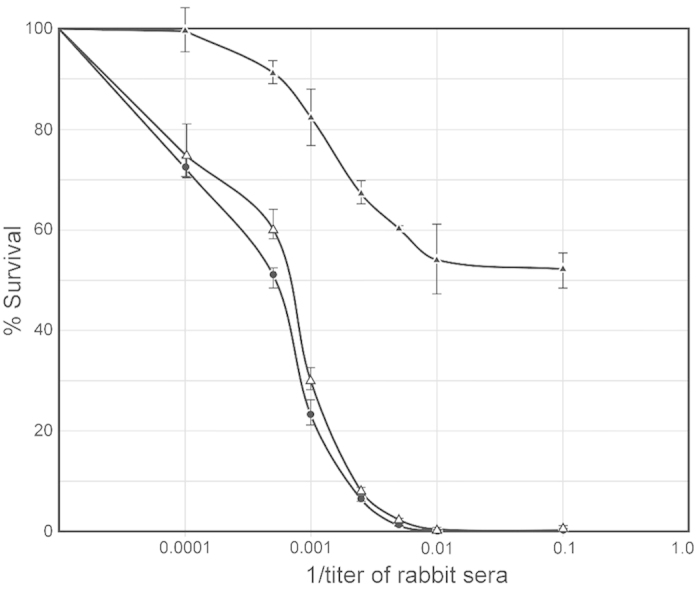
All Neisseria gonorrhoeae strains whose DNA sequences have been determined possess filamentous phage DNA sequences. To ascertain if phage encoded proteins could form the basis of a gonococcal vaccine, rabbits were orally infected with S. enterica Typhimurium strain χ3987 harboring phagemid NgoΦ6 fm. The elicited sera contained large quantities of anti-phage IgG and IgA antibodies that bound to the surface of N. gonorrhoeae cells, as shown by indirect fluorescent analysis and flow cytometry. The elicited sera was able to bind to several phage proteins. The sera also had bactericidal activity. These data demonstrate that N. gonorrhoeae filamentous phage can induce antibodies with anti-gonococcal activity and that phage proteins may be a candidate for vaccine development.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

Similar articles
-
Phage proteins are expressed on the surface of Neisseria gonorrhoeae and are potential vaccine candidates.PLoS One. 2018 Aug 23;13(8):e0202437. doi: 10.1371/journal.pone.0202437. eCollection 2018. PLoS One. 2018. PMID: 30138416 Free PMC article.
-
Association of host proteins with the broad host range filamentous phage NgoΦ6 of Neisseria gonorrhoeae.PLoS One. 2020 Oct 15;15(10):e0240579. doi: 10.1371/journal.pone.0240579. eCollection 2020. PLoS One. 2020. PMID: 33057372 Free PMC article.
-
Immunization with recombinant truncated Neisseria meningitidis-Macrophage Infectivity Potentiator (rT-Nm-MIP) protein induces murine antibodies that are cross-reactive and bactericidal for Neisseria gonorrhoeae.Vaccine. 2018 Jun 22;36(27):3926-3936. doi: 10.1016/j.vaccine.2018.05.069. Epub 2018 May 24. Vaccine. 2018. PMID: 29803329 Free PMC article.
-
Strategies for mimicking Neisserial saccharide epitopes as vaccines.Int Rev Immunol. 2001;20(2):229-50. doi: 10.3109/08830180109043036. Int Rev Immunol. 2001. PMID: 11878767 Review.
-
Progress Toward a Gonococcal Vaccine: The Way Forward.Front Immunol. 2019 Oct 15;10:2417. doi: 10.3389/fimmu.2019.02417. eCollection 2019. Front Immunol. 2019. PMID: 31681305 Free PMC article. Review.
Cited by
-
A New Vaccination Method Based on Phage NgoΦ6 and Its Phagemid Derivatives.Front Microbiol. 2022 Apr 28;13:793205. doi: 10.3389/fmicb.2022.793205. eCollection 2022. Front Microbiol. 2022. PMID: 35572628 Free PMC article.
-
Bacterial Vaccine Antigen Discovery in the Reverse Vaccinology 2.0 Era: Progress and Challenges.Front Immunol. 2018 Oct 8;9:2315. doi: 10.3389/fimmu.2018.02315. eCollection 2018. Front Immunol. 2018. PMID: 30349542 Free PMC article. Review.
-
Phage proteins are expressed on the surface of Neisseria gonorrhoeae and are potential vaccine candidates.PLoS One. 2018 Aug 23;13(8):e0202437. doi: 10.1371/journal.pone.0202437. eCollection 2018. PLoS One. 2018. PMID: 30138416 Free PMC article.
-
Addressing Sexually Transmitted Infections Due to Neisseria gonorrhoeae in the Present and Future.Microorganisms. 2024 Apr 28;12(5):884. doi: 10.3390/microorganisms12050884. Microorganisms. 2024. PMID: 38792714 Free PMC article. Review.
-
The Optimal Management of Neisseria gonorrhoeae Infections.Microorganisms. 2022 Dec 1;10(12):2388. doi: 10.3390/microorganisms10122388. Microorganisms. 2022. PMID: 36557641 Free PMC article. Review.
References
-
- Macaluso M. et al. A public health focus on infertility prevention, detection, and management. Fertil. Steril. 93, 16 e11–10 (2010). - PubMed
-
- Kasper D. L., Rice P. A. & McCormick W. M. Bactericidal antibody in genital infection due to Neisseria gonorrhoeae. J. Infect. Dis. 135, 243–251 (1977). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
