

Habitat-Associated Life History and Stress-Tolerance Variation in Arabidopsis arenosa
- PMID: 26941193
- PMCID: PMC4854687
- DOI: 10.1104/pp.15.01875
Habitat-Associated Life History and Stress-Tolerance Variation in Arabidopsis arenosa
Abstract
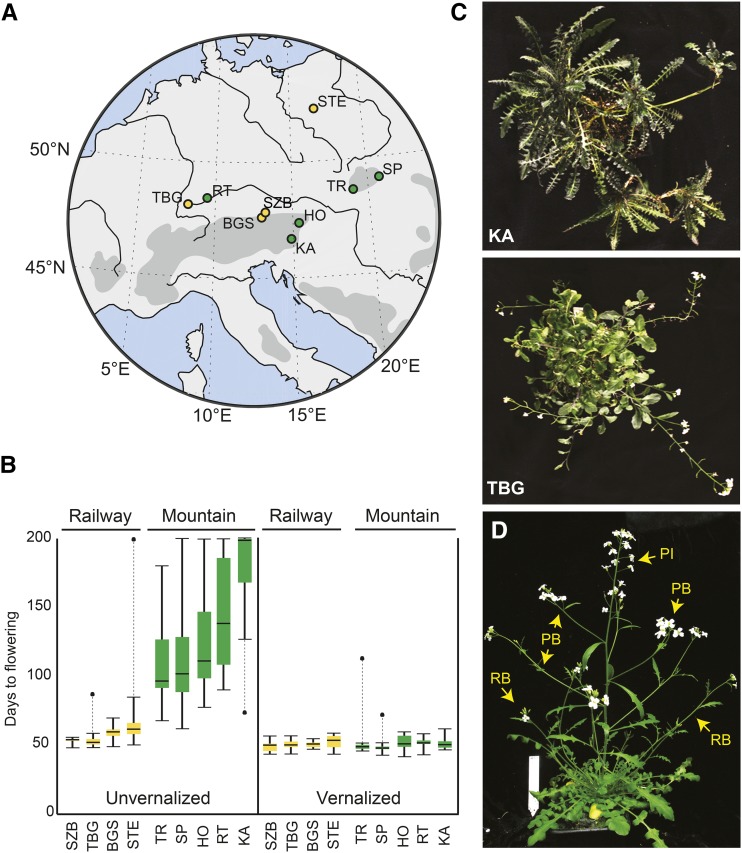
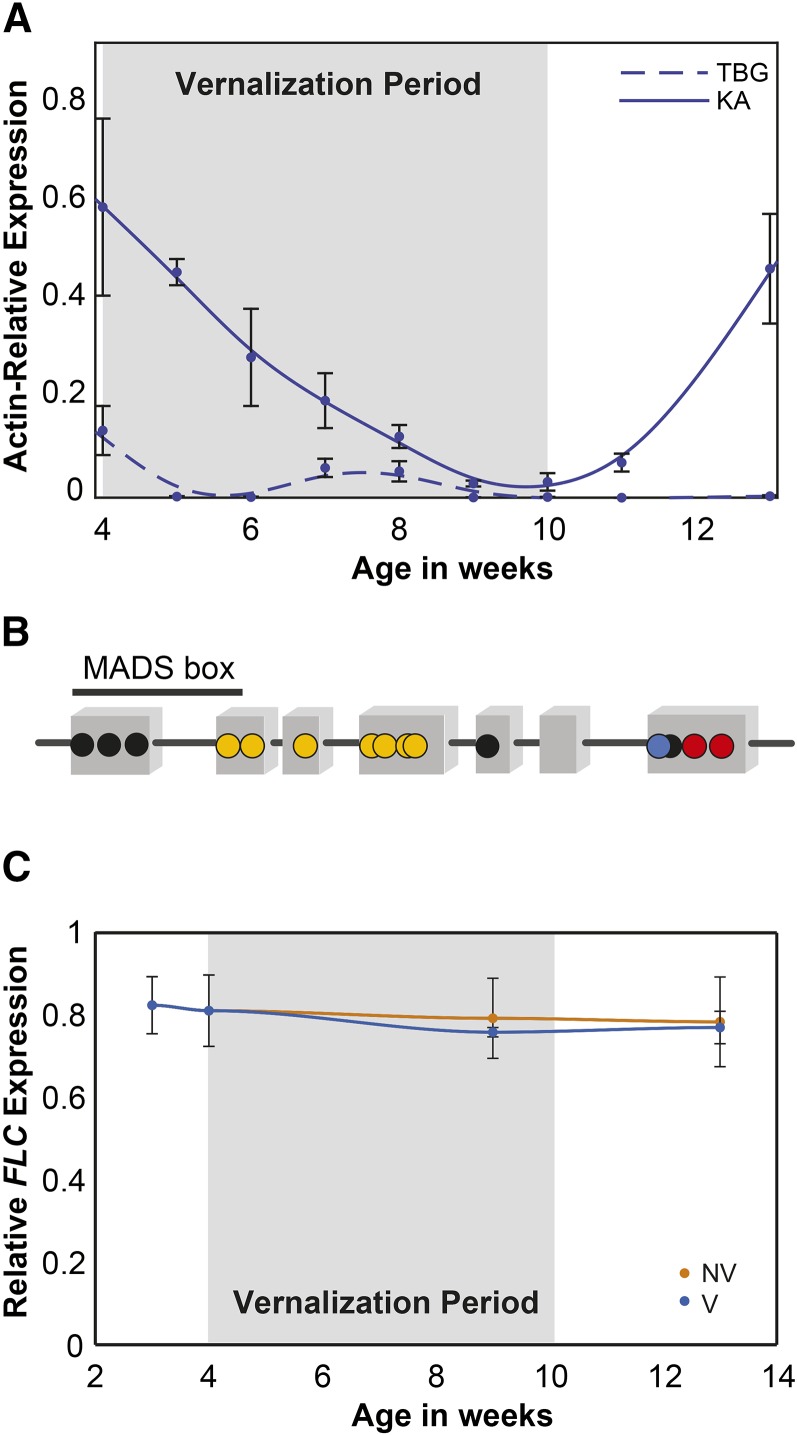
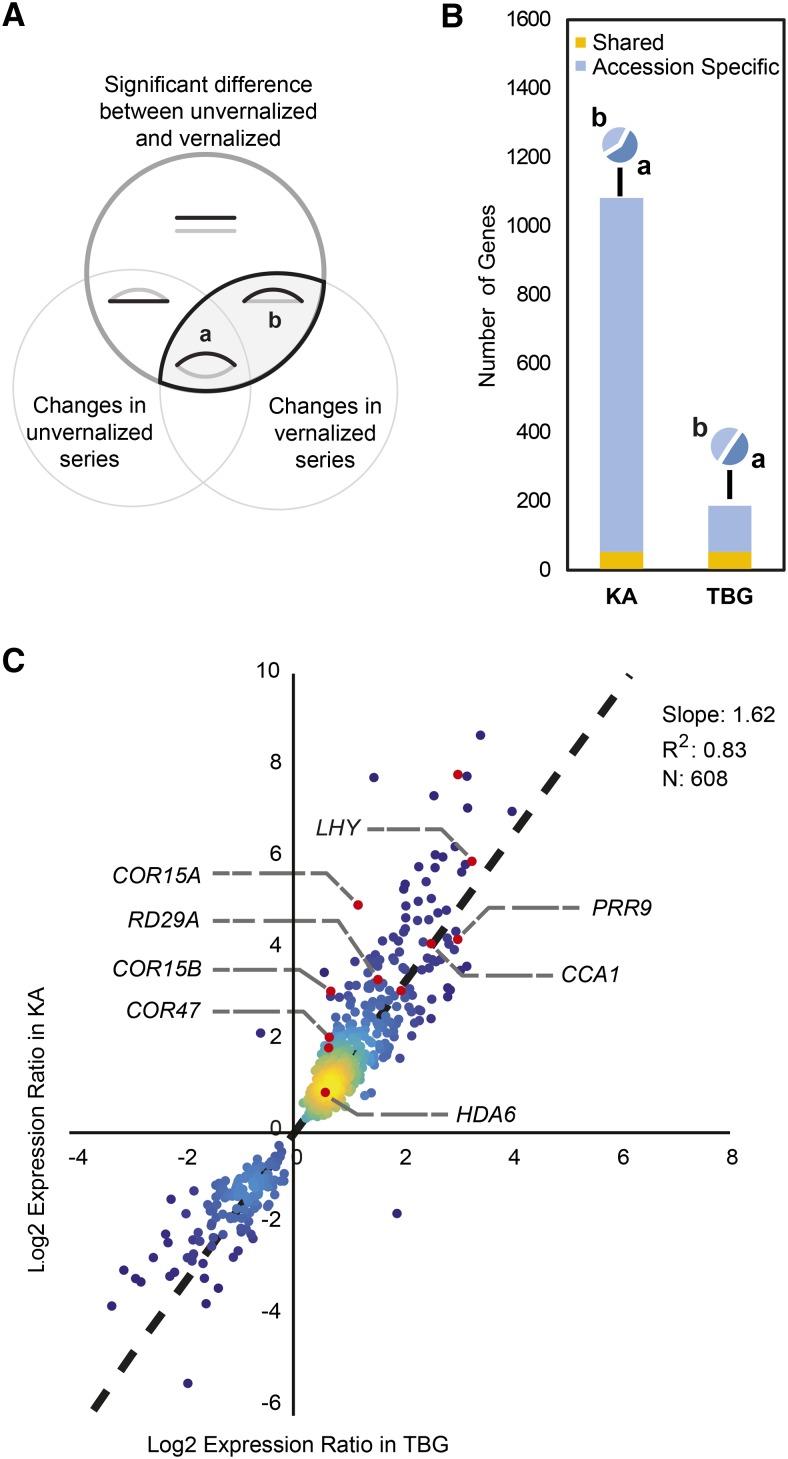
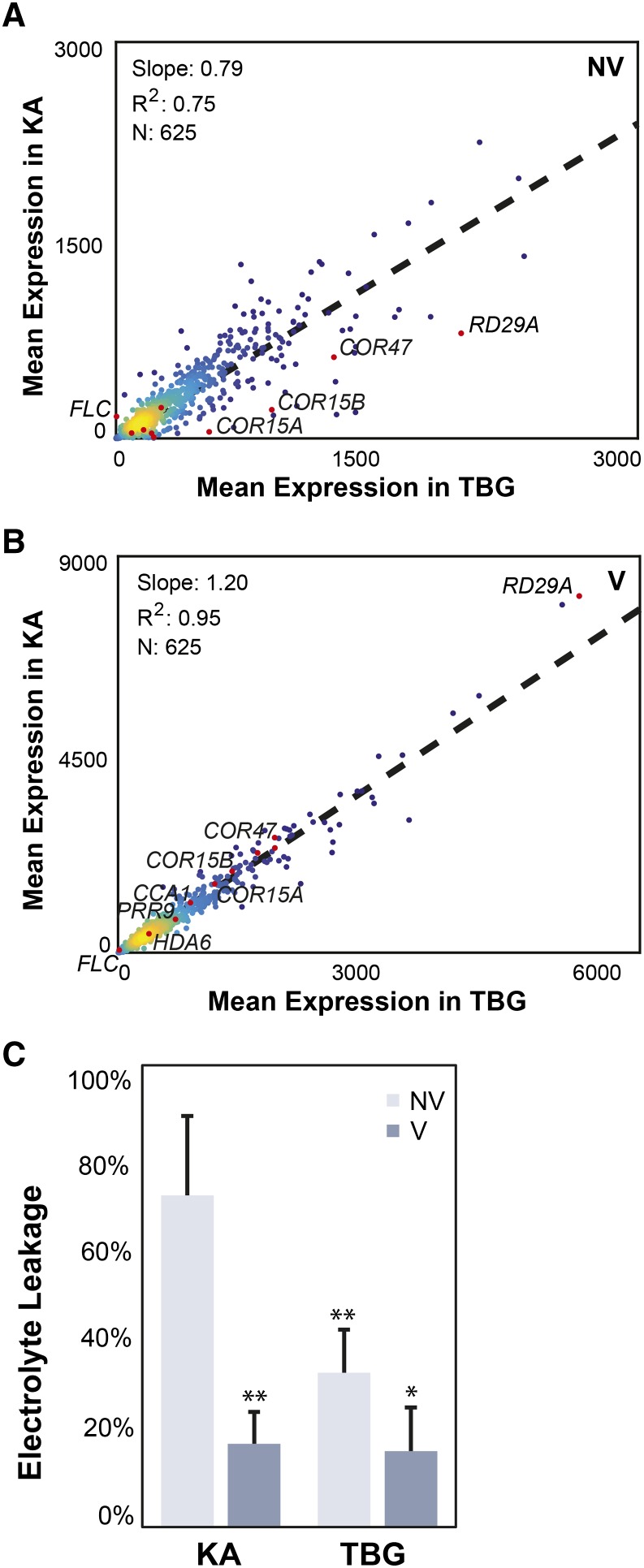
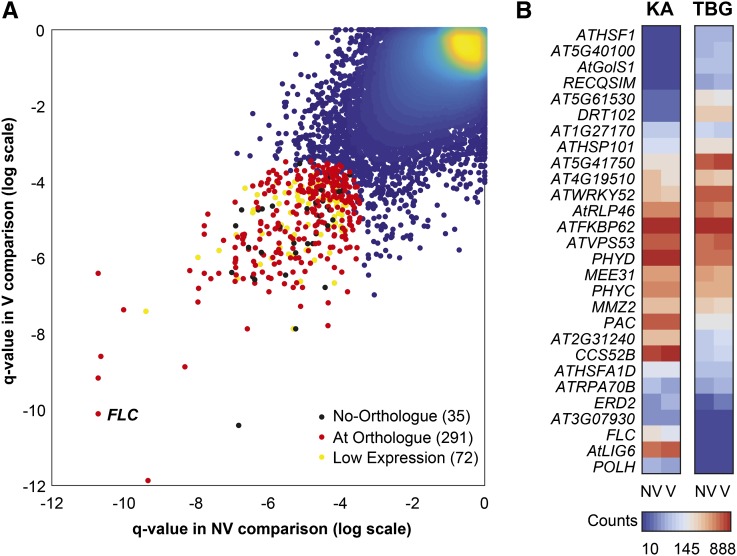
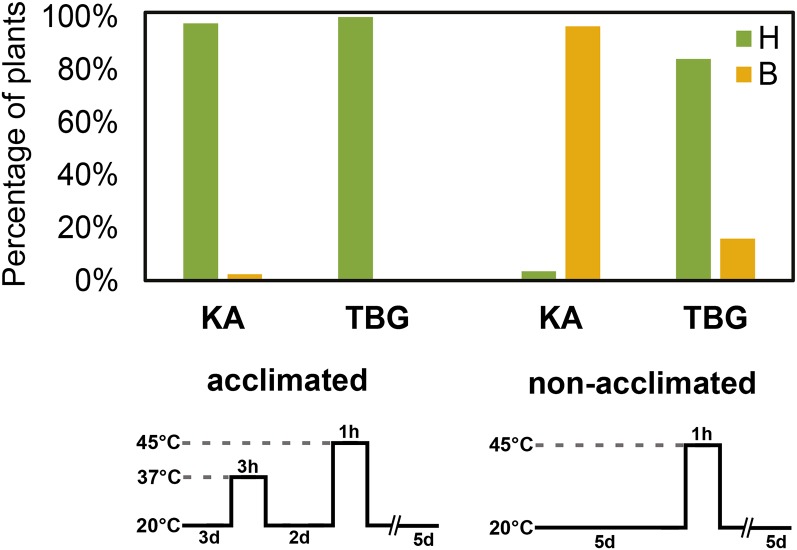
Weediness in ephemeral plants is commonly characterized by rapid cycling, prolific all-in flowering, and loss of perenniality. Many species made transitions to weediness of this sort, which can be advantageous in high-disturbance or human-associated habitats. The molecular basis of this shift, however, remains mostly mysterious. Here, we use transcriptome sequencing, genome resequencing scans for selection, and stress tolerance assays to study a weedy population of the otherwise nonweedy Arabidopsis arenosa, an obligately outbreeding relative of Arabidopsis thaliana Although weedy A. arenosa is widespread, a single genetic lineage colonized railways throughout central and northern Europe. We show that railway plants, in contrast to plants from sheltered outcrops in hill/mountain regions, are rapid cycling, have lost the vernalization requirement, show prolific flowering, and do not return to vegetative growth. Comparing transcriptomes of railway and mountain plants across time courses with and without vernalization, we found that railway plants have sharply abrogated vernalization responsiveness and high constitutive expression of heat- and cold-responsive genes. Railway plants also have strong constitutive heat shock and freezing tolerance compared with mountain plants, where tolerance must be induced. We found 20 genes with good evidence of selection in the railway population. One of these, LATE ELONGATED HYPOCOTYL, is known in A. thaliana to regulate many stress-response genes that we found to be differentially regulated among the distinct habitats. Our data suggest that, beyond life history regulation, other traits like basal stress tolerance also are associated with the evolution of weediness in A. arenosa.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Alabadí D, Oyama T, Yanovsky MJ, Harmon FG, Más P, Kay SA (2001) Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293: 880–883 - PubMed
-
- Anderson HJ, Vonarx EJ, Pastushok L, Nakagawa M, Katafuchi A, Gruz P, Di Rubbo A, Grice DM, Osmond MJ, Sakamoto AN, et al. (2008) Arabidopsis thaliana Y-family DNA polymerase eta catalyses translesion synthesis and interacts functionally with PCNA2. Plant J 55: 895–908 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
