

Beyond the Medial Regions of Prefrontal Cortex in the Regulation of Fear and Anxiety
- PMID: 26941618
- PMCID: PMC4761915
- DOI: 10.3389/fnsys.2016.00012
Beyond the Medial Regions of Prefrontal Cortex in the Regulation of Fear and Anxiety
Abstract
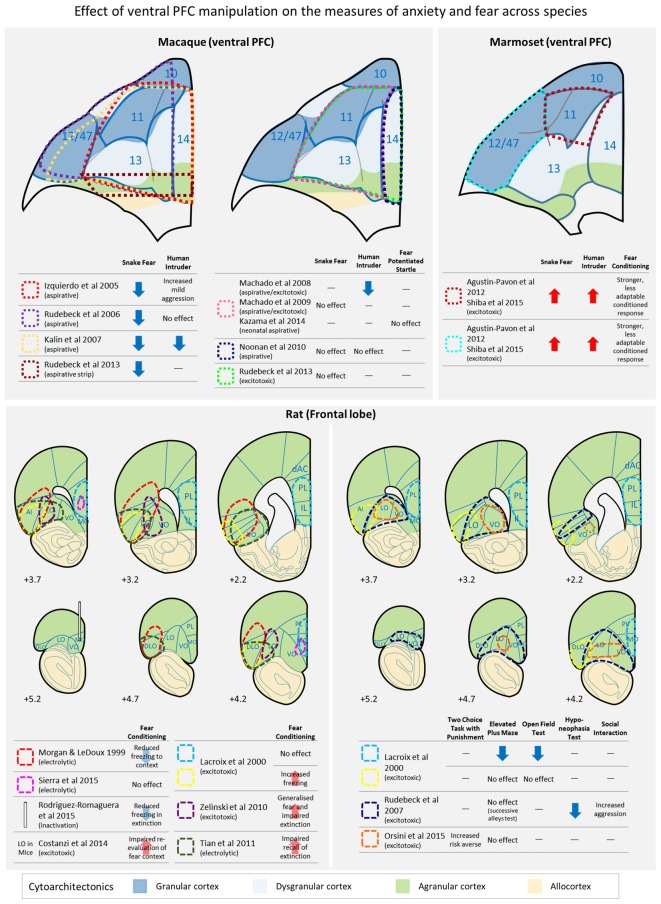
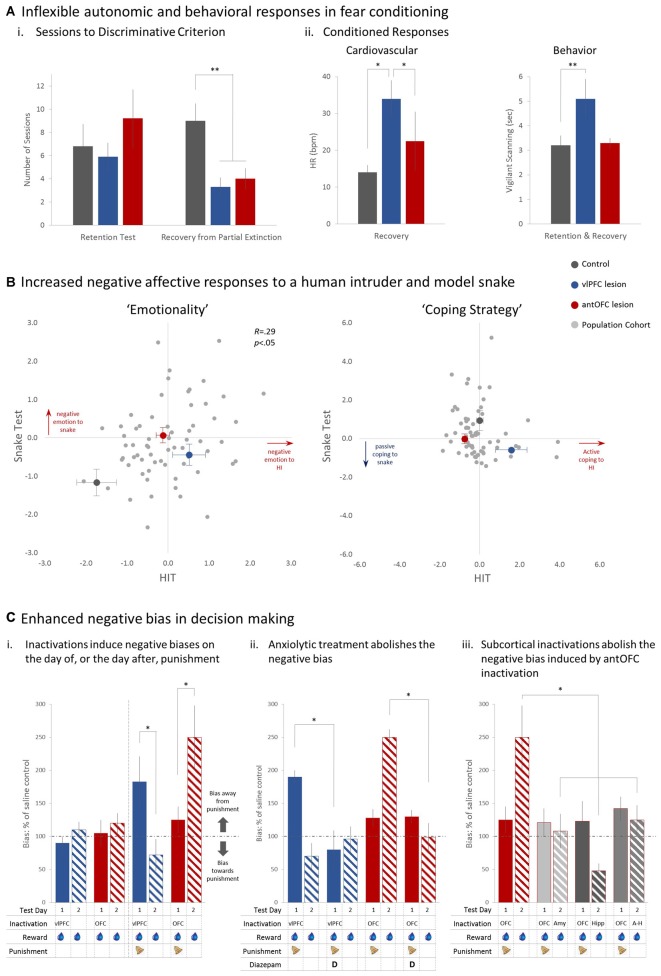
Fear and anxiety are adaptive responses but if left unregulated, or inappropriately regulated, they become biologically and socially maladaptive. Dysregulated emotions are manifest in a wide variety of psychiatric and neurological conditions but the external expression gives little indication of the underlying causes, which are inevitably multi-determined. To go beyond the overt phenotype and begin to understand the causal mechanisms leading to conditions characterized by anxiety and disorders of mood, it is necessary to identify the base psychological processes that have become dysregulated, and map them on to their associated neural substrates. So far, attention has been focused primarily on the medial regions of prefrontal cortex (PFC) and in particular their contribution to the expression and extinction of conditioned fear. However, functional neuroimaging studies have shown that the sphere of influence within the PFC is not restricted to its medial regions, but extends into dorsal, ventrolateral (vlPFC) and orbitofrontal (OFC) regions too; although the causal role of these other areas in the regulation of fear and anxiety remains to be determined and in the case of the OFC, existing findings are conflicting. Here, we review the evidence for the contribution of these other regions in negative emotion regulation in rodents and old world and new world monkeys. We consider a variety of different contexts, including conditioned and innate fear, learned and unlearned anxiety and cost-benefit decision-making, and a range of physiological and behavioral measures of emotion. It is proposed that both the OFC and vlPFC contribute to emotion regulation via their involvement, respectively, in the prediction of future outcomes and higher-order attentional control. The fractionation of these neurocognitive and neurobehavioral systems that regulate fear and anxiety opens up new opportunities for diagnostic stratification and personalized treatment strategies.
Keywords: anxiety; emotion regulation; fear; marmoset; orbitofrontal cortex; prefrontal cortex; primate; ventrolateral prefrontal cortex.
Figures
Similar articles
-
Lesions of either anterior orbitofrontal cortex or ventrolateral prefrontal cortex in marmoset monkeys heighten innate fear and attenuate active coping behaviors to predator threat.Front Syst Neurosci. 2015 Jan 21;8:250. doi: 10.3389/fnsys.2014.00250. eCollection 2014. Front Syst Neurosci. 2015. PMID: 25653599 Free PMC article.
-
Lesions of ventrolateral prefrontal or anterior orbitofrontal cortex in primates heighten negative emotion.Biol Psychiatry. 2012 Aug 15;72(4):266-72. doi: 10.1016/j.biopsych.2012.03.007. Epub 2012 Apr 12. Biol Psychiatry. 2012. PMID: 22502990
-
Converging Prefronto-Insula-Amygdala Pathways in Negative Emotion Regulation in Marmoset Monkeys.Biol Psychiatry. 2017 Dec 15;82(12):895-903. doi: 10.1016/j.biopsych.2017.06.016. Epub 2017 Jun 23. Biol Psychiatry. 2017. PMID: 28756869 Free PMC article.
-
Prefrontal Regulation of Threat-Elicited Behaviors: A Pathway to Translation.Annu Rev Psychol. 2020 Jan 4;71:357-387. doi: 10.1146/annurev-psych-010419-050905. Epub 2019 Oct 17. Annu Rev Psychol. 2020. PMID: 31622562 Review.
-
The Role of the Medial Prefrontal Cortex in the Conditioning and Extinction of Fear.Front Behav Neurosci. 2015 Nov 9;9:298. doi: 10.3389/fnbeh.2015.00298. eCollection 2015. Front Behav Neurosci. 2015. PMID: 26617500 Free PMC article. Review.
Cited by
-
Impact of Stress on Brain Morphology: Insights into Structural Biomarkers of Stress-related Disorders.Curr Neuropharmacol. 2024;22(5):935-962. doi: 10.2174/1570159X21666230703091435. Curr Neuropharmacol. 2024. PMID: 37403395 Free PMC article. Review.
-
The resilient brain: psychological resilience mediates the effect of amplitude of low-frequency fluctuations in orbitofrontal cortex on subjective well-being in young healthy adults.Soc Cogn Affect Neurosci. 2018 Sep 4;13(7):755-763. doi: 10.1093/scan/nsy045. Soc Cogn Affect Neurosci. 2018. PMID: 29939335 Free PMC article.
-
Silica nanoparticles induce neurodegeneration-like changes in behavior, neuropathology, and affect synapse through MAPK activation.Part Fibre Toxicol. 2018 Jul 3;15(1):28. doi: 10.1186/s12989-018-0263-3. Part Fibre Toxicol. 2018. PMID: 29970116 Free PMC article.
-
Expatriates' Multiple Fears, from Terrorism to Working Conditions: Development of a Model.Front Psychol. 2016 Oct 13;7:1571. doi: 10.3389/fpsyg.2016.01571. eCollection 2016. Front Psychol. 2016. PMID: 27790173 Free PMC article.
-
Multimodal Imaging Analysis Reveals Frontal-Associated Networks in Relation to Individual Resilience Strength.Int J Environ Res Public Health. 2021 Jan 27;18(3):1123. doi: 10.3390/ijerph18031123. Int J Environ Res Public Health. 2021. PMID: 33513995 Free PMC article.
References
-
- Baxter M. G., Gaffan D., Kyriazis D. A., Mitchell A. S. (2009). Ventrolateral prefrontal cortex is required for performance of a strategy implementation task but not reinforcer devaluation effects in rhesus monkeys. Eur. J. Neurosci. 29, 2049–2059. 10.1111/j.1460-9568.2009.06740.x - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous