

Arabidopsis thaliana: A Model Host Plant to Study Plant-Pathogen Interaction Using Rice False Smut Isolates of Ustilaginoidea virens
- PMID: 26941759
- PMCID: PMC4763110
- DOI: 10.3389/fpls.2016.00192
Arabidopsis thaliana: A Model Host Plant to Study Plant-Pathogen Interaction Using Rice False Smut Isolates of Ustilaginoidea virens
Abstract
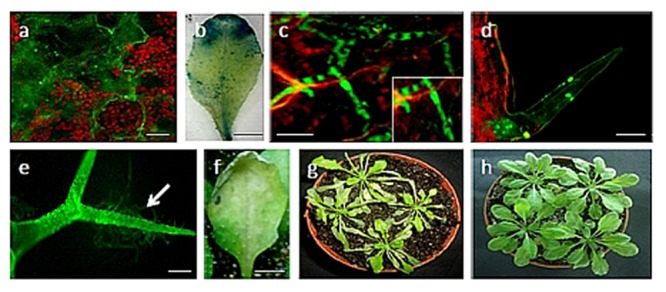
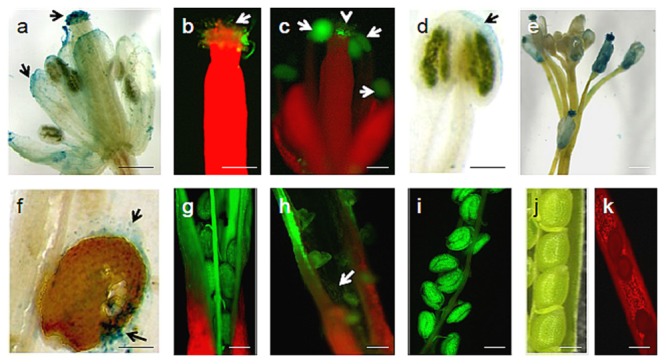
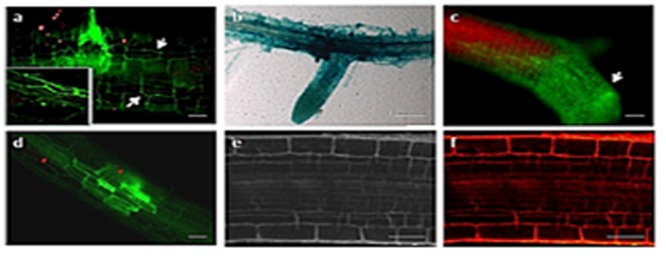
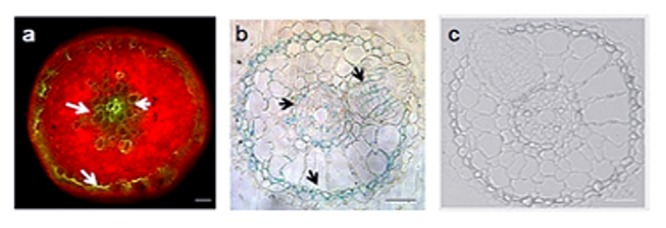
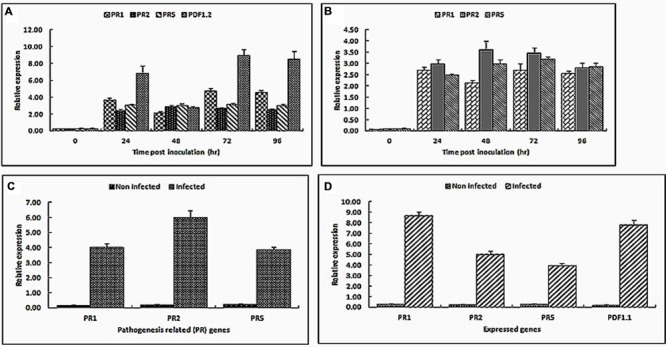
Rice false smut fungus which is a biotrophic fungal pathogen causes an important rice disease and brings a severe damage where rice is cultivated. We established a new fungal-plant pathosystem where Ustilaginoidea virens was able to interact compatibly with the model plant Arabidopsis thaliana. Disease symptoms were apparent on the leaves of the plants after 6 days of post inoculation in the form of chlorosis. Cytological studies showed that U. virens caused a heavy infestation inside the cells of the chlorotic tissues. Development and colonization of aerial mycelia in association with floral organ, particularly on anther and stigma of the flowers after 3 weeks of post inoculation was evident which finally caused infection on the developing seeds and pod tissues. The fungus adopts a uniquely biotrophic infection strategy in roots and spreads without causing a loss of host cell viability. We have also demonstrated that U. virens isolates infect Arabidopsis and the plant subsequently activates different defense response mechanisms which are witnessed by the expression of pathogenesis-related genes, PR-1, PR-2, PR-5, PDF1.1, and PDF1.2. The established A. thaliana-U. virens pathosystem will now permit various follow-up molecular genetics and gene expression experiments to be performed to identify the defense signals and responses that restrict fungal hyphae colonization in planta and also provide initial evidence for tissue-adapted fungal infection strategies.
Keywords: Arabidopsis thaliana; PR genes; Ustilaginoidea virens; endophytic colonization; plant defensin gene; plant–pathogen interaction.
Figures

References
-
- Asano Y., Hiramoto T., Nishino R., Aiba Y., Kimura T., Yoshihara K., et al. (2012). Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. Gastrointest. Liver Physiol. 303 G1288–G1295. 10.1152/ajpgi.00341.2012 - DOI - PubMed
-
- Bohman S., Staal J., Thomma B. P., Wang M., Dixelius C. (2004). Characterisation of an Arabidopsis-Leptosphaeria maculans pathosystem: resistance partially requires camalexin biosynthesis and is independent of salicylic acid, ethylene and jasmonic acid signalling. Plant J. 37 9–20. 10.1046/j.1365-313X.2003.01927.x - DOI - PubMed
-
- Chamberlain A. C. (1975). “The movement of plant particles in plant communities,” in Vegetation and the Atmosphere ed. Monteith J. L. (New York, NY: Academic Press; ) 155–204.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials