

The Mitochondrial Basis of Aging
- PMID: 26942670
- PMCID: PMC4779179
- DOI: 10.1016/j.molcel.2016.01.028
The Mitochondrial Basis of Aging
Abstract
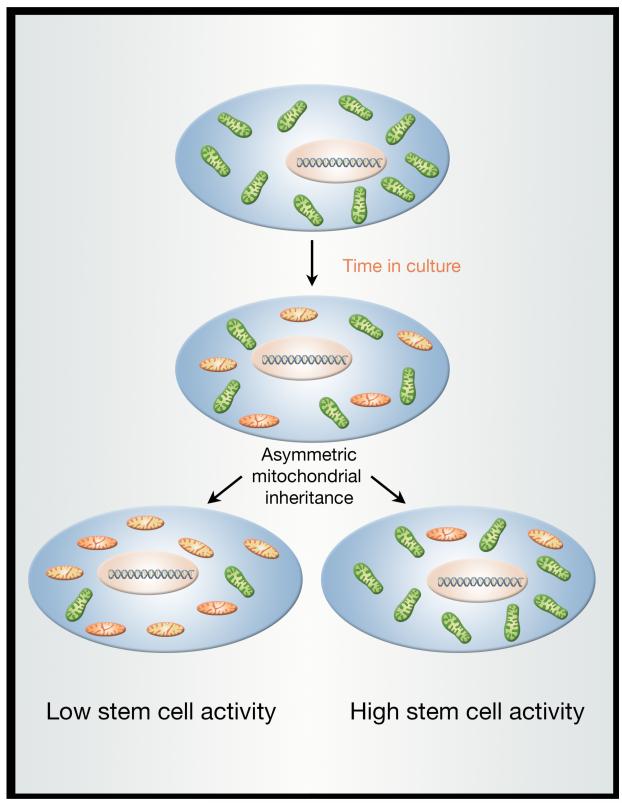
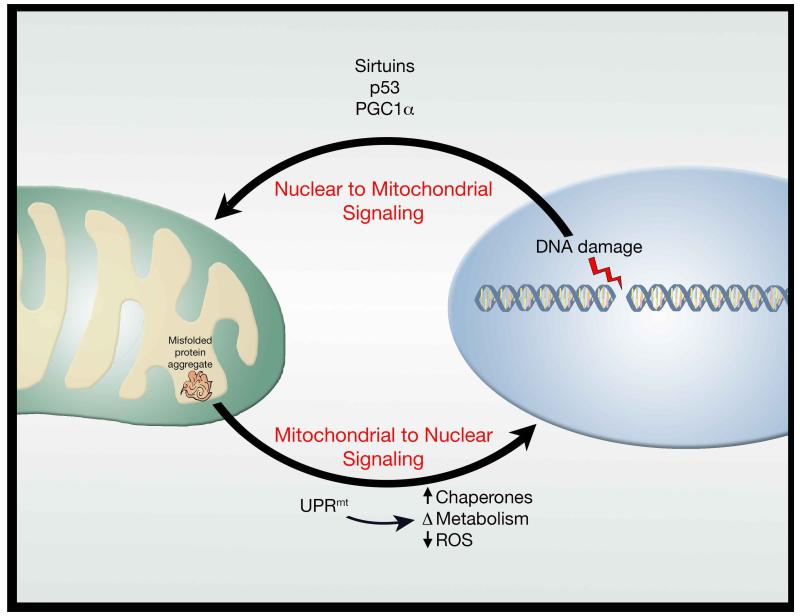
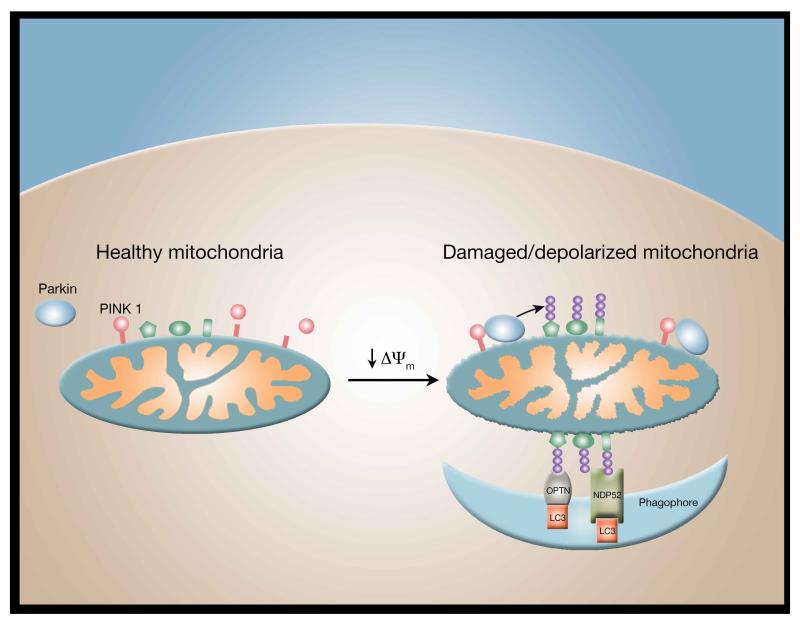
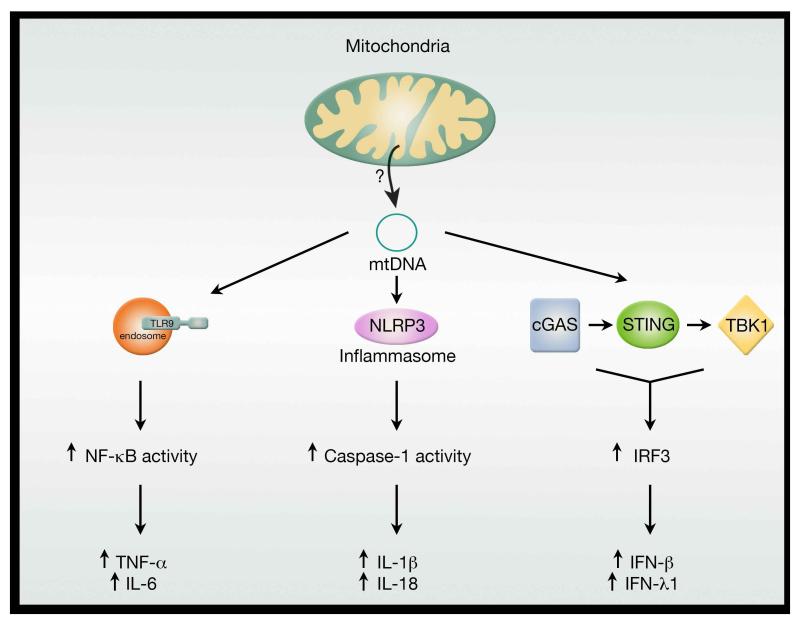
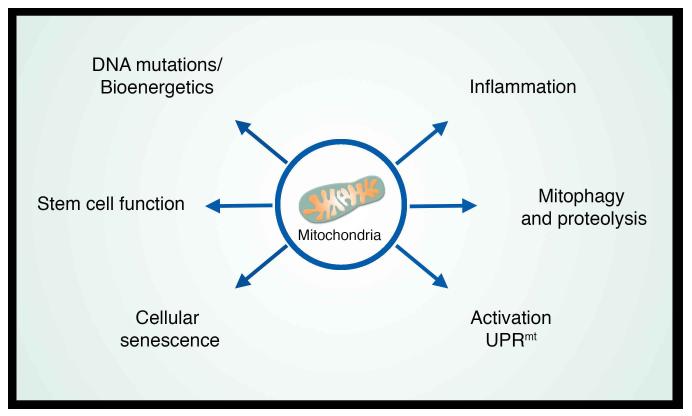
A decline in mitochondrial quality and activity has been associated with normal aging and correlated with the development of a wide range of age-related diseases. Here, we review the evidence that a decline in mitochondria function contributes to aging. In particular, we discuss how mitochondria contribute to specific aspects of the aging process, including cellular senescence, chronic inflammation, and the age-dependent decline in stem cell activity. Signaling pathways regulating the mitochondrial unfolded protein response and mitophagy are also reviewed, with particular emphasis placed on how these pathways might, in turn, regulate longevity. Taken together, these observations suggest that mitochondria influence or regulate a number of key aspects of aging and suggest that strategies directed at improving mitochondrial quality and function might have far-reaching beneficial effects.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Ahlqvist KJ, Hamalainen RH, Yatsuga S, Uutela M, Terzioglu M, Gotz A, Forsstrom S, Salven P, Angers-Loustau A, Kopra OH, et al. Somatic progenitor cell vulnerability to mitochondrial DNA mutagenesis underlies progeroid phenotypes in Polg mutator mice. Cell Metab. 2012;15:100–109. - PubMed
-
- Arany Z, He H, Lin J, Hoyer K, Handschin C, Toka O, Ahmad F, Matsui T, Chin S, Wu PH, et al. Transcriptional coactivator PGC-1 alpha controls the energy state and contractile function of cardiac muscle. Cell Metab. 2005;1:259–271. - PubMed
-
- Argiles JM, Busquets S, Stemmler B, Lopez-Soriano FJ. Cachexia and sarcopenia: mechanisms and potential targets for intervention. Curr Opin Pharmacol. 2015;22:100–106. - PubMed
-
- Austin S, St-Pierre J. PGC1alpha and mitochondrial metabolism--emerging concepts and relevance in ageing and neurodegenerative disorders. J Cell Sci. 2012;125:4963–4971. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
