

Pin1 is required for sustained B cell proliferation upon oncogenic activation of Myc
- PMID: 26943576
- PMCID: PMC5008323
- DOI: 10.18632/oncotarget.7846
Pin1 is required for sustained B cell proliferation upon oncogenic activation of Myc
Abstract
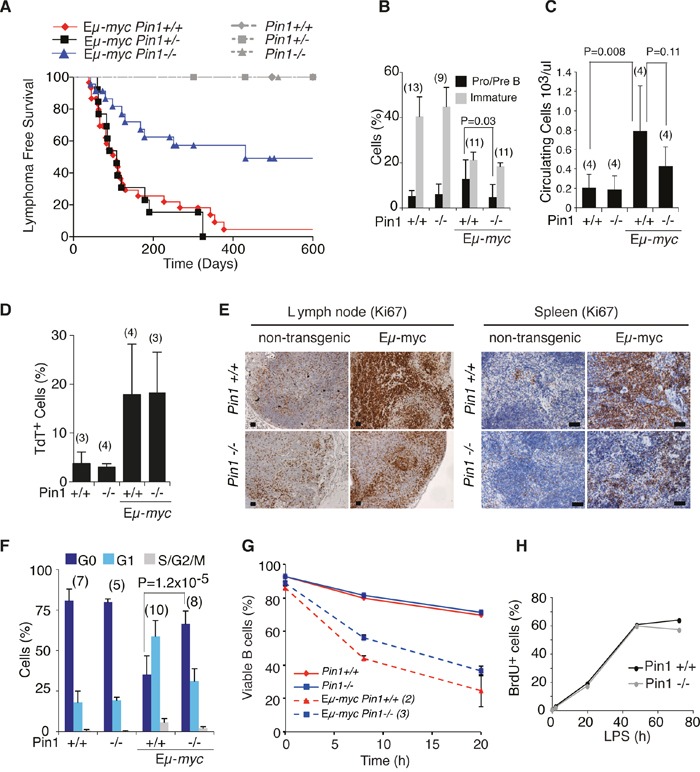
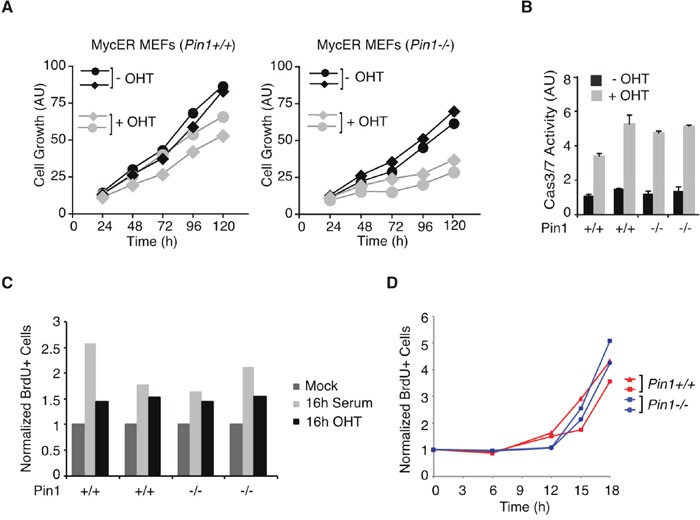
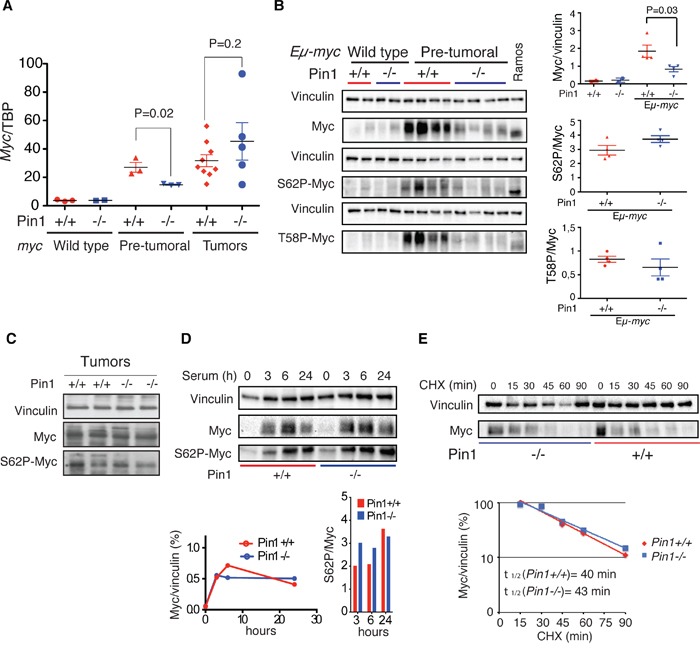
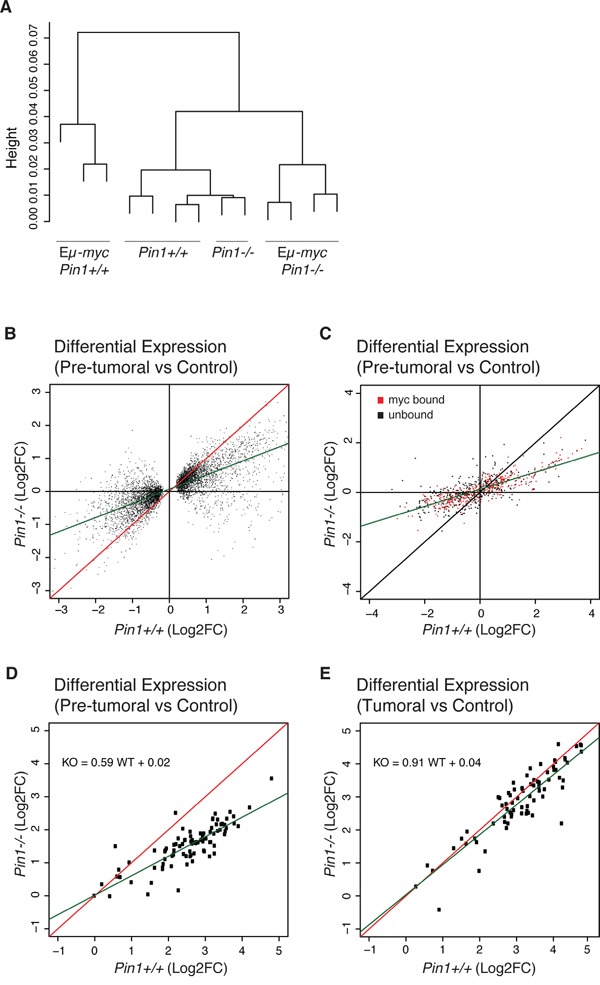
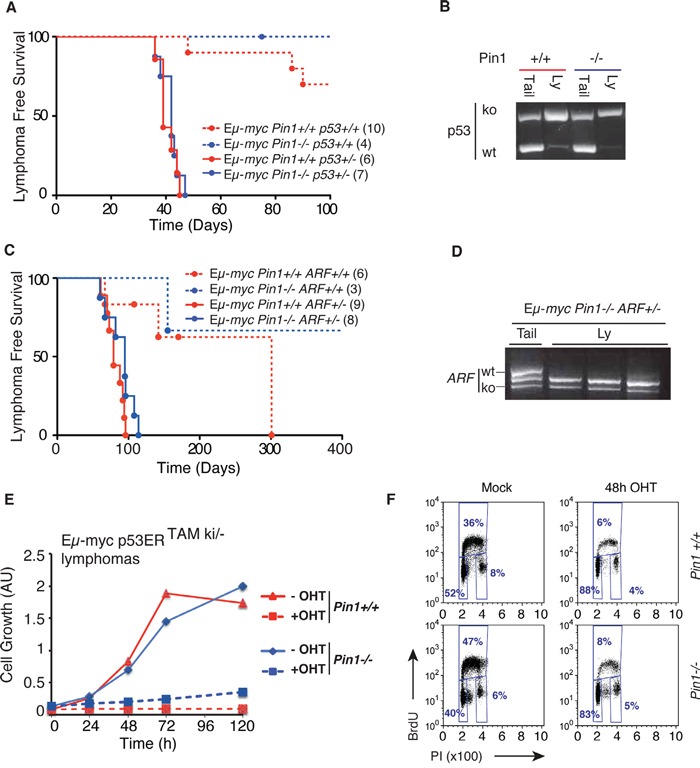
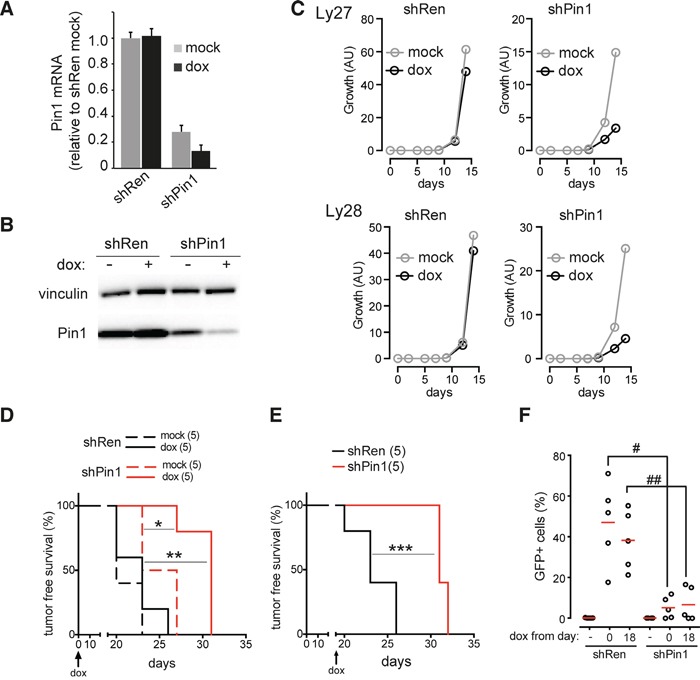
The c-myc proto-oncogene is activated by translocation in Burkitt's lymphoma and substitutions in codon 58 stabilize the Myc protein or augment its oncogenic potential. In wild-type Myc, phosphorylation of Ser 62 and Thr 58 provides a landing pad for the peptidyl prolyl-isomerase Pin1, which in turn promotes Ser 62 dephosphorylation and Myc degradation. However, the role of Pin1 in Myc-induced lymphomagenesis remains unknown. We show here that genetic ablation of Pin1 reduces lymphomagenesis in Eμ-myc transgenic mice. In both Pin1-deficient B-cells and MEFs, the proliferative response to oncogenic Myc was selectively impaired, with no alterations in Myc-induced apoptosis or mitogen-induced cell cycle entry. This proliferative defect wasn't attributable to alterations in either Ser 62 phosphorylation or Myc-regulated transcription, but instead relied on the activity of the ARF-p53 pathway. Pin1 silencing in lymphomas retarded disease progression in mice, making Pin1 an attractive therapeutic target in Myc-driven tumors.
Keywords: Pin1; c-myc; lymphoma; proliferation.
Conflict of interest statement
The Authors declare no competing financial interests.
Figures
References
-
- Lu KP, Hanes SD, Hunter T. A human peptidyl-prolyl isomerase essential for regulation of mitosis. Nature. 1996;380:544–547. - PubMed
-
- Yaffe MB, Schutkowski M, Shen M, Zhou XZ, Stukenberg PT, Rahfeld JU, Xu J, Kuang J, Kirschner MW, Fischer G, Cantley LC, Lu KP. Sequence-specific and phosphorylation-dependent proline isomerization: a potential mitotic regulatory mechanism. Science. 1997;278:1957–1960. - PubMed
-
- Ranganathan R, Lu KP, Hunter T, Noel JP. Structural and functional analysis of the mitotic rotamase Pin1 suggests substrate recognition is phosphorylation dependent. Cell. 1997;89:875–886. - PubMed
-
- Lu KP, Zhou XZ. The prolyl isomerase PIN1: a pivotal new twist in phosphorylation signalling and disease. Nat Rev Mol Cell Biol. 2007;8:904–916. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
