

Thalamocortical Projections onto Behaviorally Relevant Neurons Exhibit Plasticity during Adult Motor Learning
- PMID: 26948893
- PMCID: PMC4795975
- DOI: 10.1016/j.neuron.2016.02.001
Thalamocortical Projections onto Behaviorally Relevant Neurons Exhibit Plasticity during Adult Motor Learning
Abstract
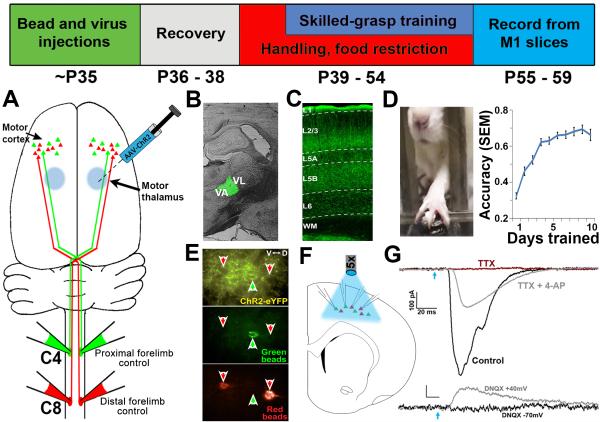
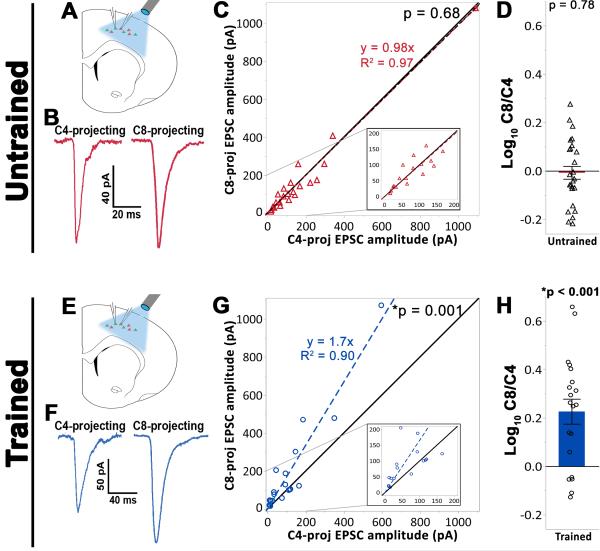
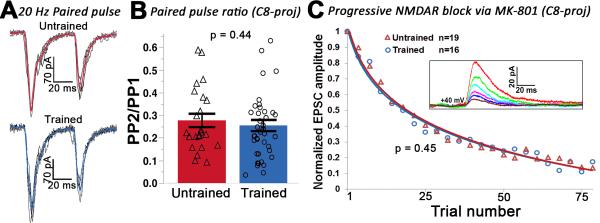
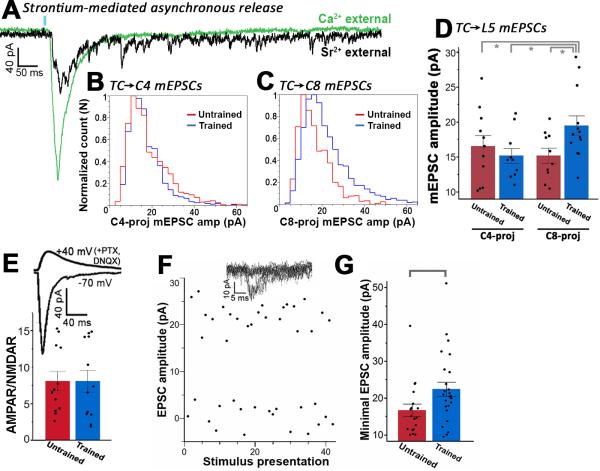
Layer 5 neurons of the neocortex receive direct and relatively strong input from the thalamus. However, the intralaminar distribution of these inputs and their capacity for plasticity in adult animals are largely unknown. In slices of the primary motor cortex (M1), we simultaneously recorded from pairs of corticospinal neurons associated with control of distinct motor outputs: distal forelimb versus proximal forelimb. Activation of ChR2-expressing thalamocortical afferents in M1 before motor learning produced equivalent responses in monosynaptic excitation of neurons controlling the distal and proximal forelimb, suggesting balanced thalamic input at baseline. Following skilled grasp training, however, thalamocortical input shifted to bias activation of corticospinal neurons associated with control of the distal forelimb. This increase was associated with a cell-specific increase in mEPSC amplitude but not presynaptic release probability. These findings demonstrate distinct and highly segregated plasticity of thalamocortical projections during adult learning.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Selective plasticity of layer 2/3 inputs onto distal forelimb controlling layer 5 corticospinal neurons with skilled grasp motor training.Cell Rep. 2024 Apr 23;43(4):113986. doi: 10.1016/j.celrep.2024.113986. Epub 2024 Apr 9. Cell Rep. 2024. PMID: 38598336
-
Dual-channel circuit mapping reveals sensorimotor convergence in the primary motor cortex.J Neurosci. 2015 Mar 11;35(10):4418-26. doi: 10.1523/JNEUROSCI.3741-14.2015. J Neurosci. 2015. PMID: 25762684 Free PMC article.
-
Strengthening of horizontal cortical connections following skill learning.Nat Neurosci. 1998 Jul;1(3):230-4. doi: 10.1038/678. Nat Neurosci. 1998. PMID: 10195148
-
Local connections of excitatory neurons in motor-associated cortical areas of the rat.Front Neural Circuits. 2013 May 28;7:75. doi: 10.3389/fncir.2013.00075. eCollection 2013. Front Neural Circuits. 2013. PMID: 23754982 Free PMC article. Review.
-
Neocortical mechanisms in motor learning.Curr Opin Neurobiol. 2003 Apr;13(2):225-31. doi: 10.1016/s0959-4388(03)00046-1. Curr Opin Neurobiol. 2003. PMID: 12744978 Review.
Cited by
-
Changes in Brain Matrix Glycan Sulfation Associate With Reactive Gliosis and Motor Coordination in Mice With Head Trauma.Front Behav Neurosci. 2021 Oct 28;15:745288. doi: 10.3389/fnbeh.2021.745288. eCollection 2021. Front Behav Neurosci. 2021. PMID: 34776892 Free PMC article.
-
Reduced thalamic excitation to motor cortical pyramidal tract neurons in parkinsonism.Sci Adv. 2023 Aug 25;9(34):eadg3038. doi: 10.1126/sciadv.adg3038. Epub 2023 Aug 23. Sci Adv. 2023. PMID: 37611096 Free PMC article.
-
Whole-brain mapping of long-range inputs to the VIP-expressing inhibitory neurons in the primary motor cortex.Front Neural Circuits. 2023 May 19;17:1093066. doi: 10.3389/fncir.2023.1093066. eCollection 2023. Front Neural Circuits. 2023. PMID: 37275468 Free PMC article.
-
Specific connectivity optimizes learning in thalamocortical loops.Cell Rep. 2024 Apr 23;43(4):114059. doi: 10.1016/j.celrep.2024.114059. Epub 2024 Apr 10. Cell Rep. 2024. PMID: 38602873 Free PMC article.
-
Rhesus macaque versus rat divergence in the corticospinal projectome.Neuron. 2022 Sep 21;110(18):2970-2983.e4. doi: 10.1016/j.neuron.2022.07.002. Epub 2022 Aug 1. Neuron. 2022. PMID: 35917818 Free PMC article.
References
-
- Akintunde A, Buxton DF. Differential sites of origin and collateralization of corticospinal neurons in the rat: a multiple fluorescent retrograde tracer study. Brain Res. 1992;575:86–92. - PubMed
