

Inhibition of fatty acid oxidation as a therapy for MYC-overexpressing triple-negative breast cancer
- PMID: 26950360
- PMCID: PMC4892846
- DOI: 10.1038/nm.4055
Inhibition of fatty acid oxidation as a therapy for MYC-overexpressing triple-negative breast cancer
Abstract
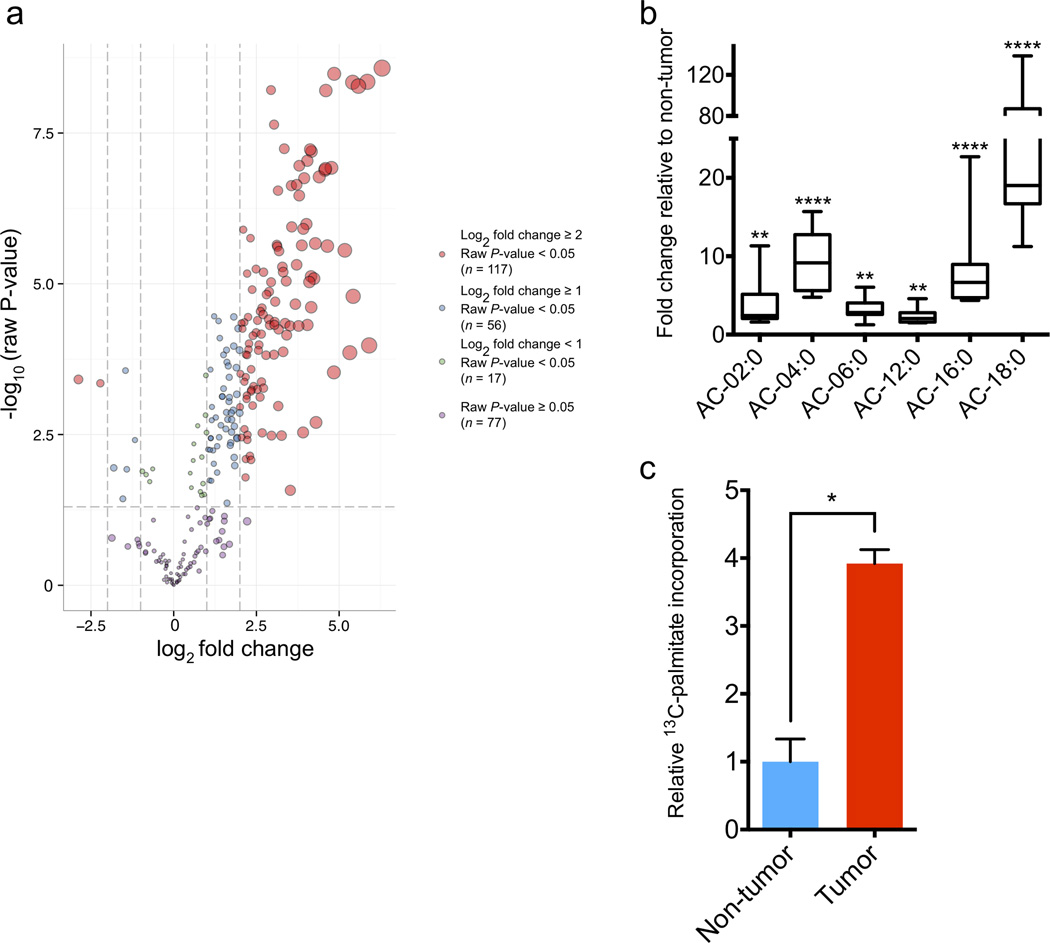
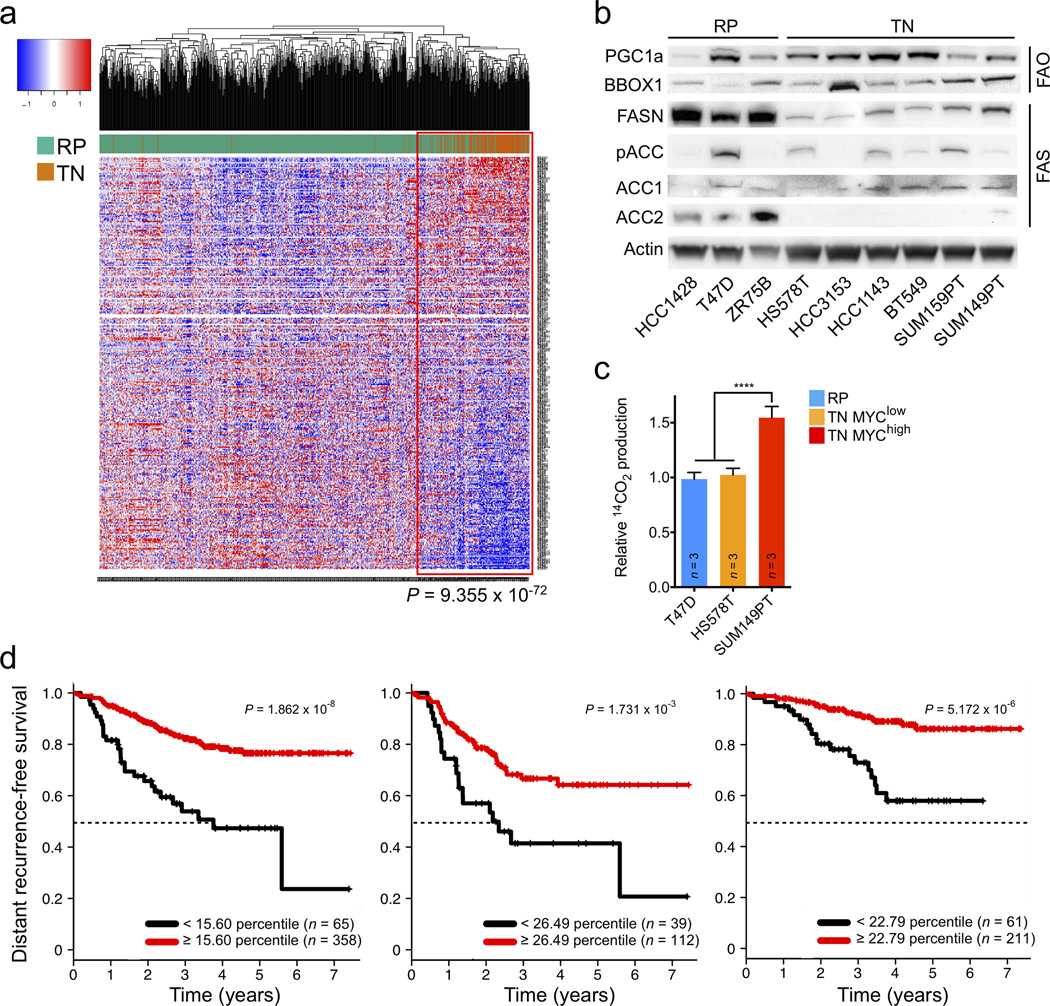
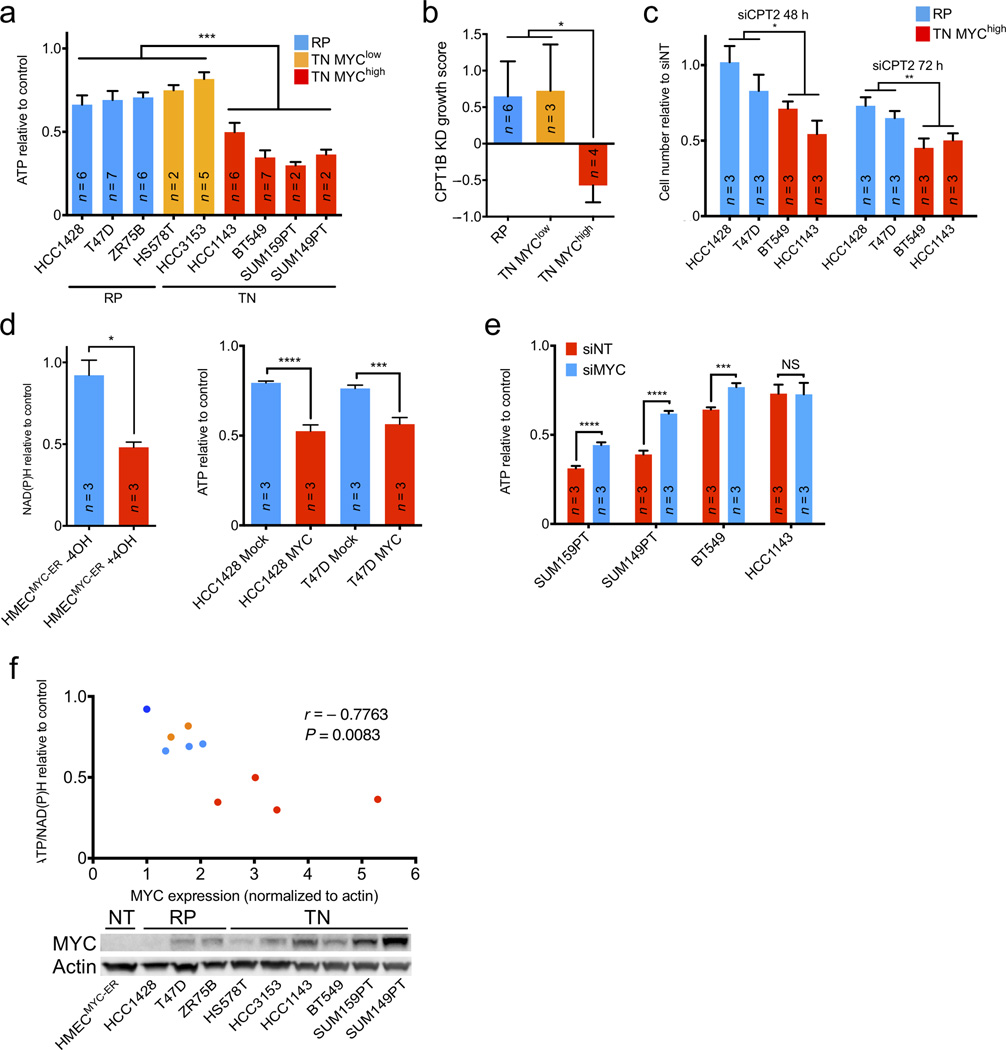
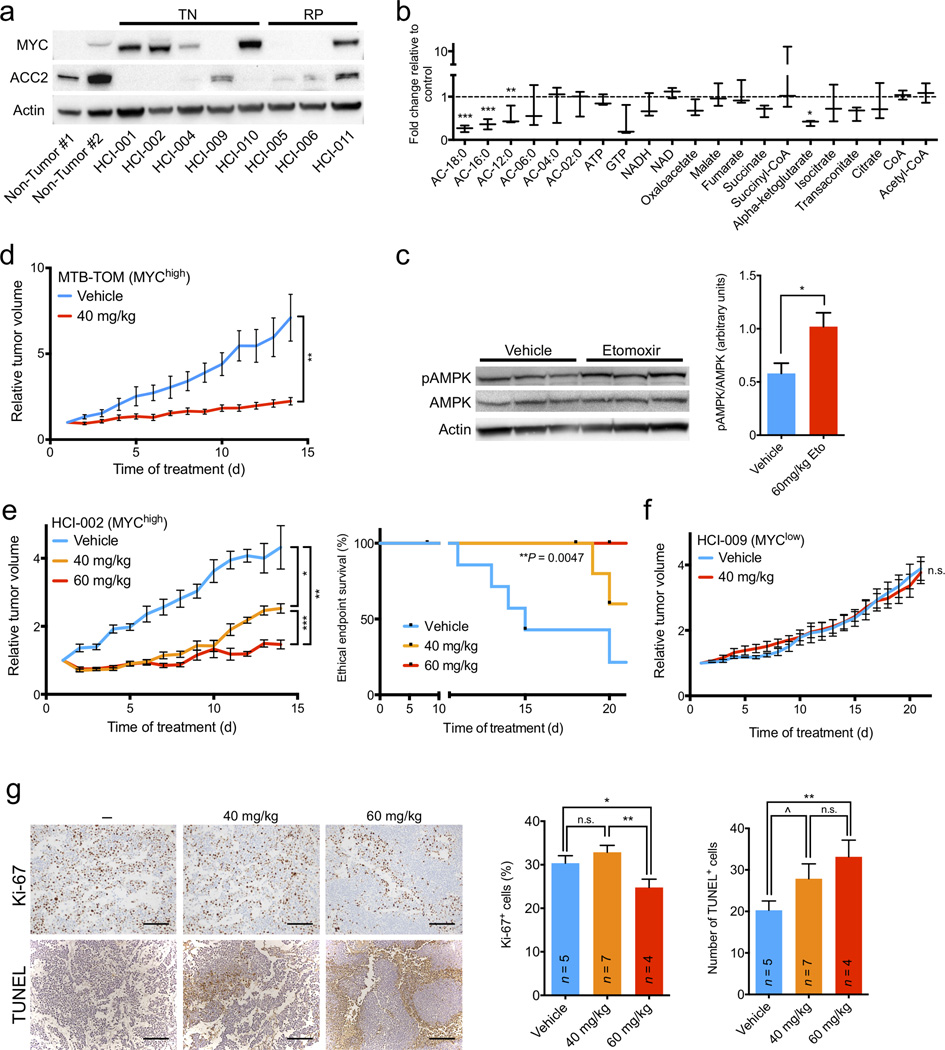
Expression of the oncogenic transcription factor MYC is disproportionately elevated in triple-negative breast cancer (TNBC), as compared to estrogen receptor-, progesterone receptor- or human epidermal growth factor 2 receptor-positive (RP) breast cancer. We and others have shown that MYC alters metabolism during tumorigenesis. However, the role of MYC in TNBC metabolism remains mostly unexplored. We hypothesized that MYC-dependent metabolic dysregulation is essential for the growth of MYC-overexpressing TNBC cells and may identify new therapeutic targets for this clinically challenging subset of breast cancer. Using a targeted metabolomics approach, we identified fatty acid oxidation (FAO) intermediates as being dramatically upregulated in a MYC-driven model of TNBC. We also identified a lipid metabolism gene signature in patients with TNBC that were identified from The Cancer Genome Atlas database and from multiple other clinical data sets, implicating FAO as a dysregulated pathway that is critical for TNBC cell metabolism. We found that pharmacologic inhibition of FAO catastrophically decreased energy metabolism in MYC-overexpressing TNBC cells and blocked tumor growth in a MYC-driven transgenic TNBC model and in a MYC-overexpressing TNBC patient-derived xenograft. These findings demonstrate that MYC-overexpressing TNBC shows an increased bioenergetic reliance on FAO and identify the inhibition of FAO as a potential therapeutic strategy for this subset of breast cancer.
Figures
Comment in
-
MYC-Overexpressing TNBCs Depend on Fatty Acid Oxidation.Cancer Discov. 2016 May;6(5):OF5. doi: 10.1158/2159-8290.CD-RW2016-049. Epub 2016 Mar 17. Cancer Discov. 2016. PMID: 26988665
References
MAIN TEXT REFERENCES
METHODS-ONLY REFERENCES
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
