

A branched-chain amino acid metabolite drives vascular fatty acid transport and causes insulin resistance
- PMID: 26950361
- PMCID: PMC4949205
- DOI: 10.1038/nm.4057
A branched-chain amino acid metabolite drives vascular fatty acid transport and causes insulin resistance
Abstract
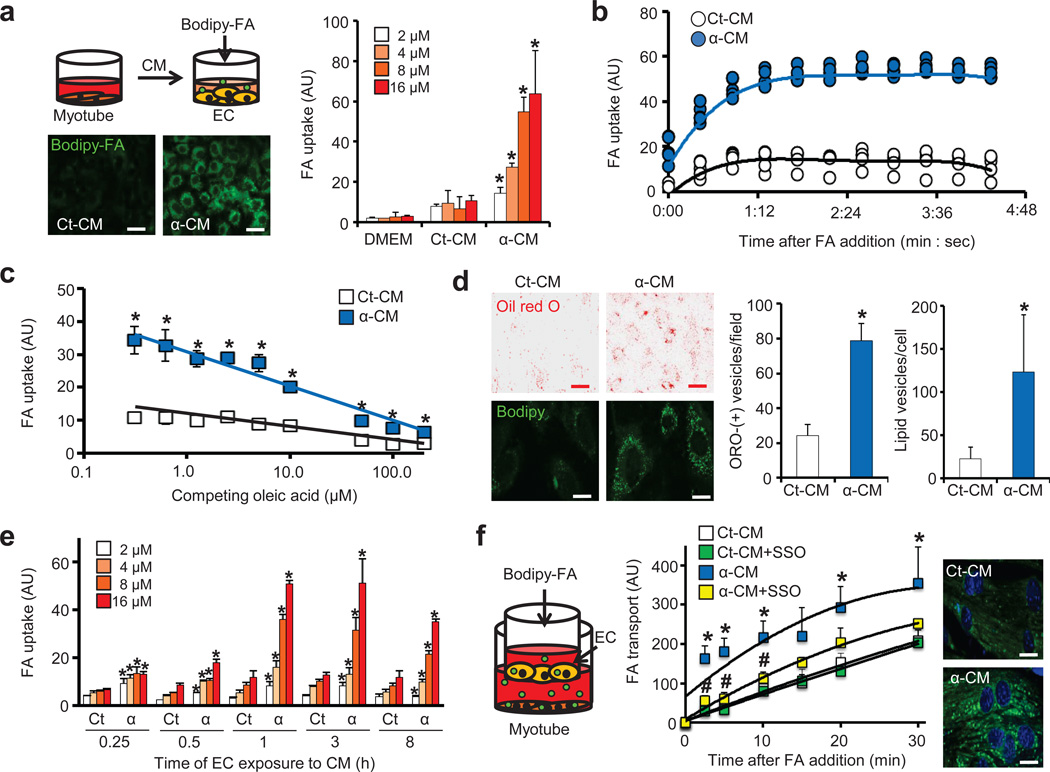
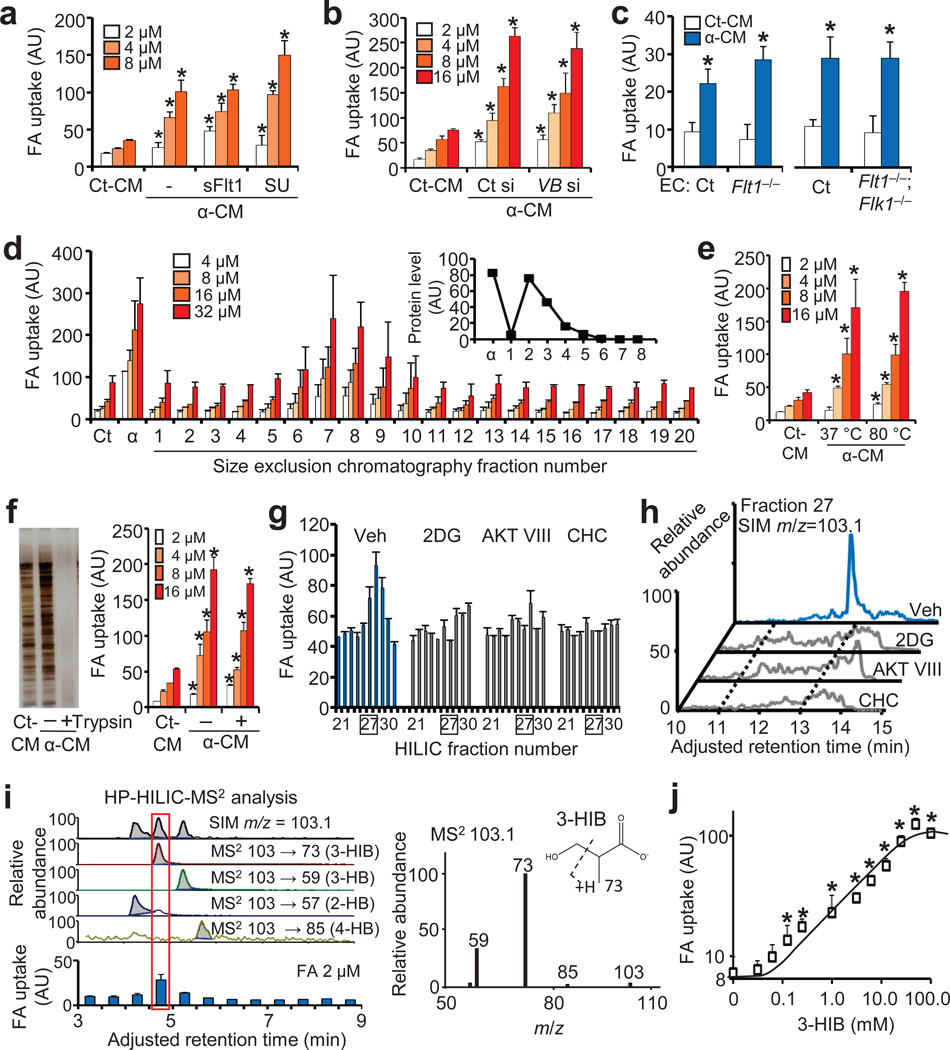
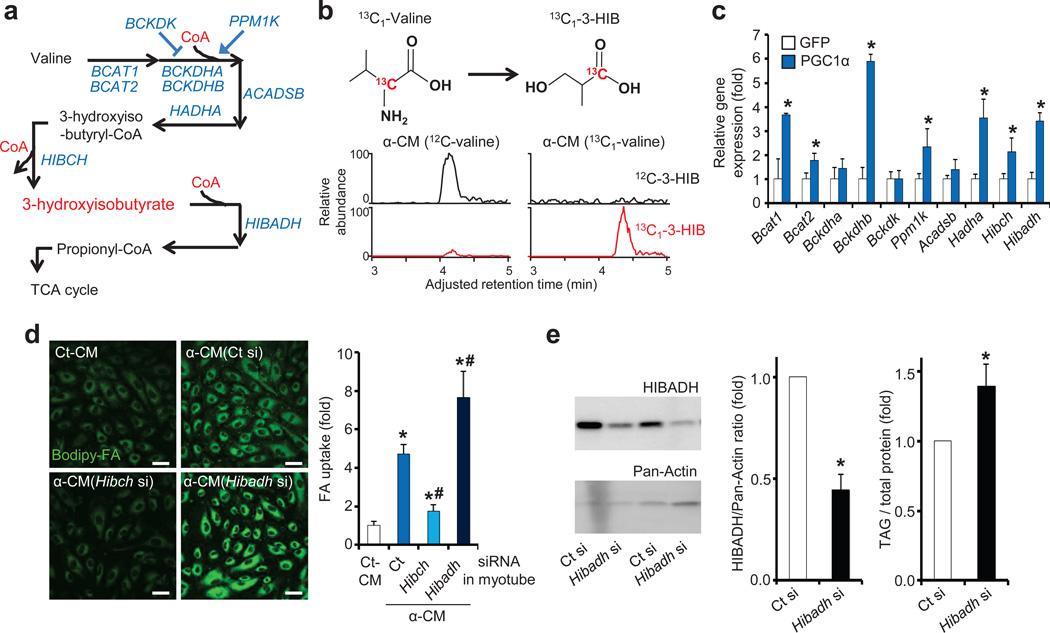
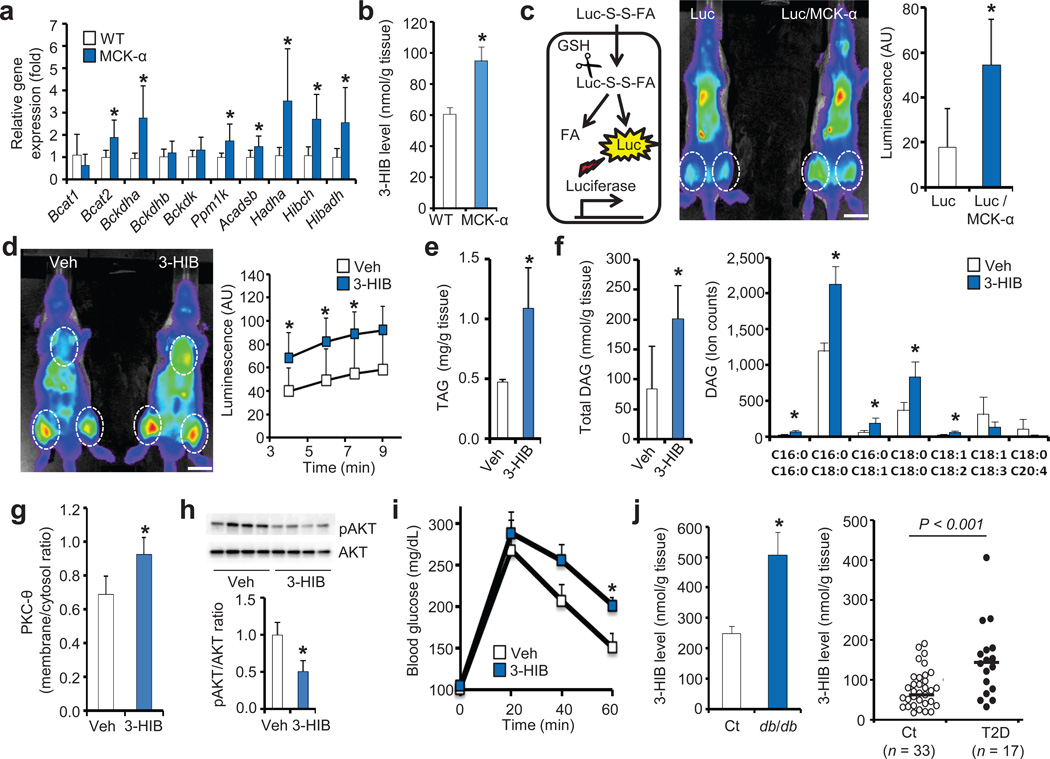
Epidemiological and experimental data implicate branched-chain amino acids (BCAAs) in the development of insulin resistance, but the mechanisms that underlie this link remain unclear. Insulin resistance in skeletal muscle stems from the excess accumulation of lipid species, a process that requires blood-borne lipids to initially traverse the blood vessel wall. How this trans-endothelial transport occurs and how it is regulated are not well understood. Here we leveraged PPARGC1a (also known as PGC-1α; encoded by Ppargc1a), a transcriptional coactivator that regulates broad programs of fatty acid consumption, to identify 3-hydroxyisobutyrate (3-HIB), a catabolic intermediate of the BCAA valine, as a new paracrine regulator of trans-endothelial fatty acid transport. We found that 3-HIB is secreted from muscle cells, activates endothelial fatty acid transport, stimulates muscle fatty acid uptake in vivo and promotes lipid accumulation in muscle, leading to insulin resistance in mice. Conversely, inhibiting the synthesis of 3-HIB in muscle cells blocks the ability of PGC-1α to promote endothelial fatty acid uptake. 3-HIB levels are elevated in muscle from db/db mice with diabetes and from human subjects with diabetes, as compared to those without diabetes. These data unveil a mechanism in which the metabolite 3-HIB, by regulating the trans-endothelial flux of fatty acids, links the regulation of fatty acid flux to BCAA catabolism, providing a mechanistic explanation for how increased BCAA catabolic flux can cause diabetes.
Figures
Comment in
-
Diabetes: New mechanism for insulin resistance.Nat Rev Endocrinol. 2016 May;12(5):249. doi: 10.1038/nrendo.2016.46. Epub 2016 Mar 18. Nat Rev Endocrinol. 2016. PMID: 26988615 No abstract available.
References
-
- Shulman GI. Ectopic fat in insulin resistance, dyslipidemia, and cardiometabolic disease. N. Engl. J. Med. 2014;371:1131–1141. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 AI007061/AI/NIAID NIH HHS/United States
- R01 DK098656/DK/NIDDK NIH HHS/United States
- R01 DK095072/DK/NIDDK NIH HHS/United States
- AR062128/AR/NIAMS NIH HHS/United States
- HL094499/HL/NHLBI NIH HHS/United States
- K01 AR062128/AR/NIAMS NIH HHS/United States
- DK098656/DK/NIDDK NIH HHS/United States
- K01 DK102771/DK/NIDDK NIH HHS/United States
- DK095072/DK/NIDDK NIH HHS/United States
- R01 HL094499/HL/NHLBI NIH HHS/United States
- R01 HL093234/HL/NHLBI NIH HHS/United States
- HL093234/HL/NHLBI NIH HHS/United States
- 5 T32 GM7592-35/GM/NIGMS NIH HHS/United States
- R01 HL125275/HL/NHLBI NIH HHS/United States
- HL125275/HL/NHLBI NIH HHS/United States
- T32 GM007592/GM/NIGMS NIH HHS/United States
- P30 DK057521/DK/NIDDK NIH HHS/United States
- P01 DK049210/DK/NIDDK NIH HHS/United States
- T32 HL007374/HL/NHLBI NIH HHS/United States
- P30 DK019525/DK/NIDDK NIH HHS/United States
- DK049210/DK/NIDDK NIH HHS/United States
- R01 AG043483/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
