

Underestimation of the Maximal Capacity of the Mitochondrial Electron Transport System in Oligomycin-Treated Cells
- PMID: 26950698
- PMCID: PMC4780810
- DOI: 10.1371/journal.pone.0150967
Underestimation of the Maximal Capacity of the Mitochondrial Electron Transport System in Oligomycin-Treated Cells
Abstract
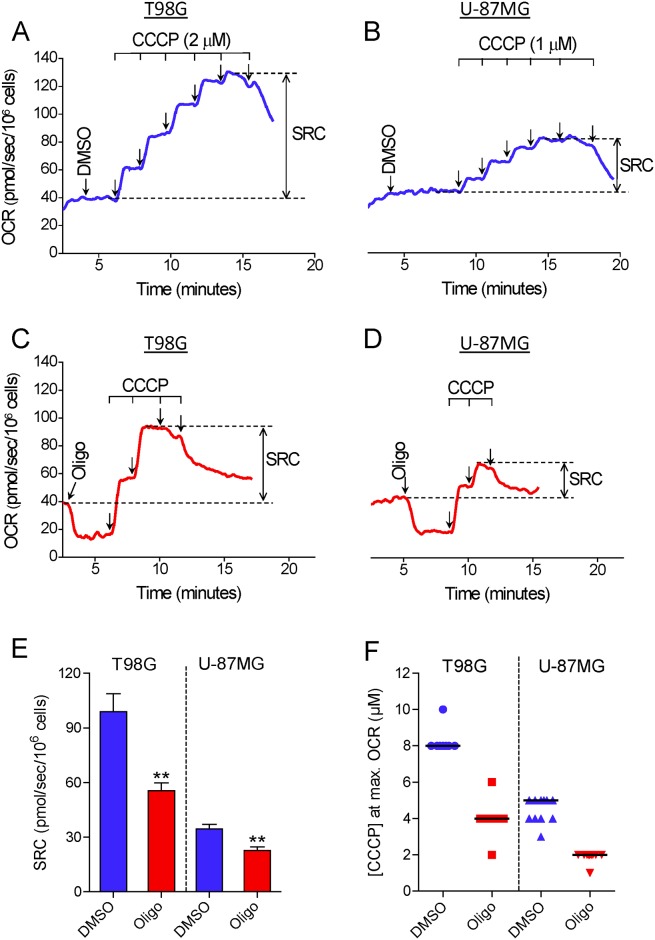
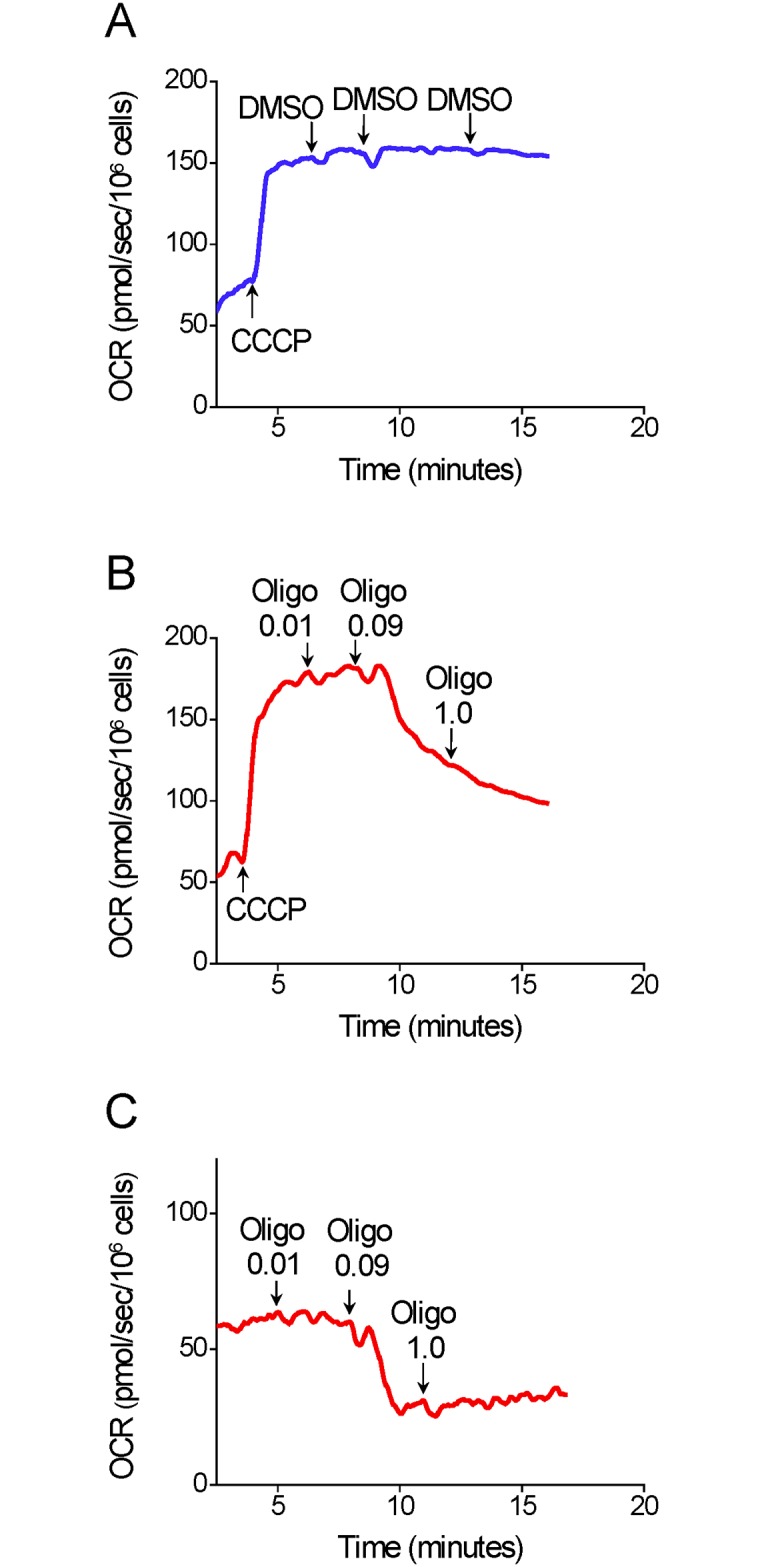
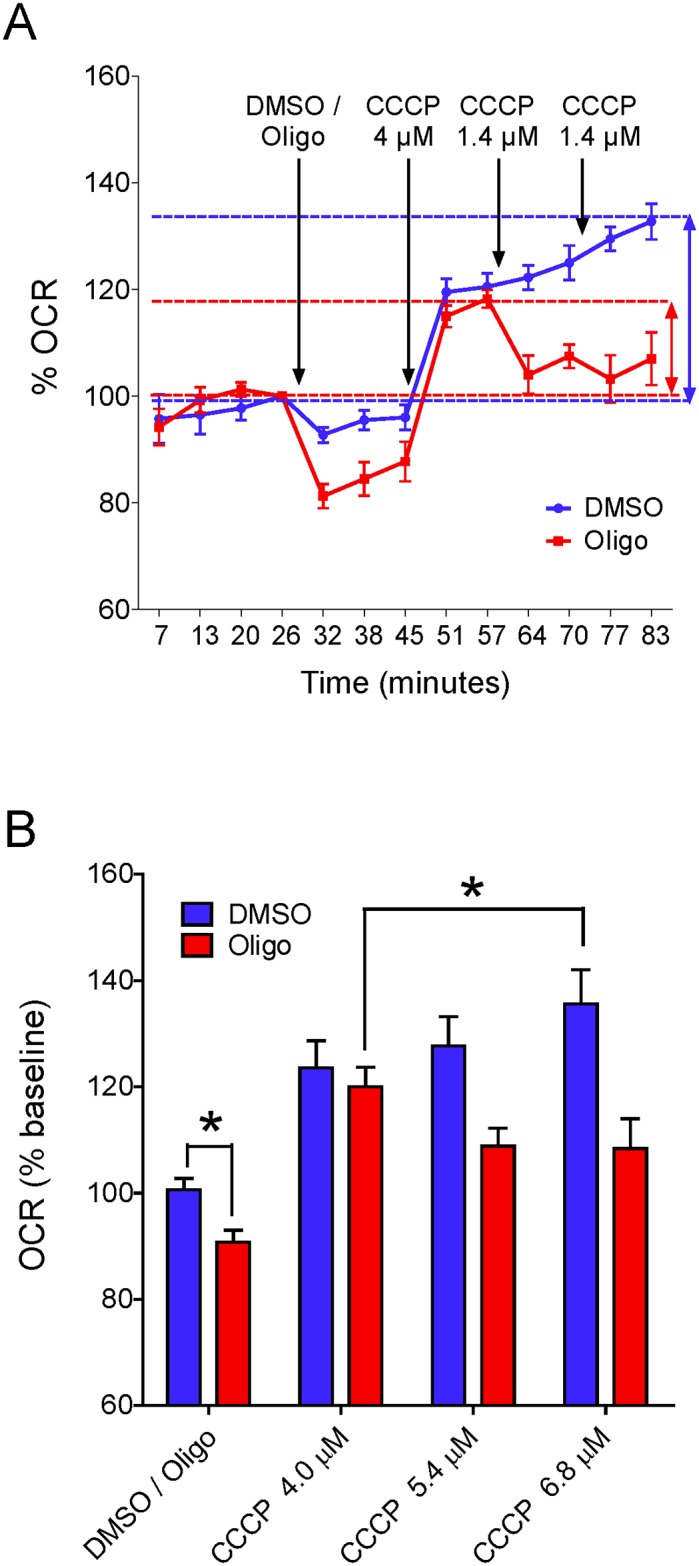
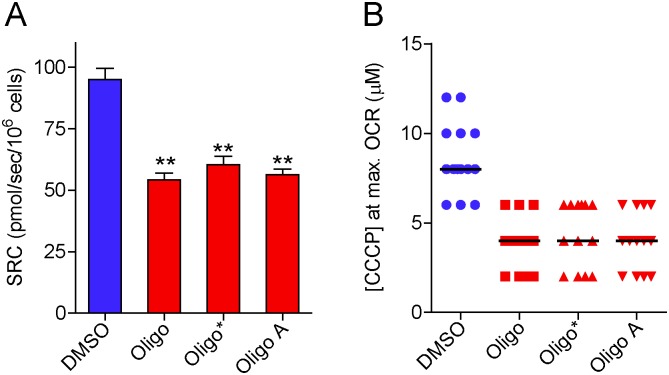
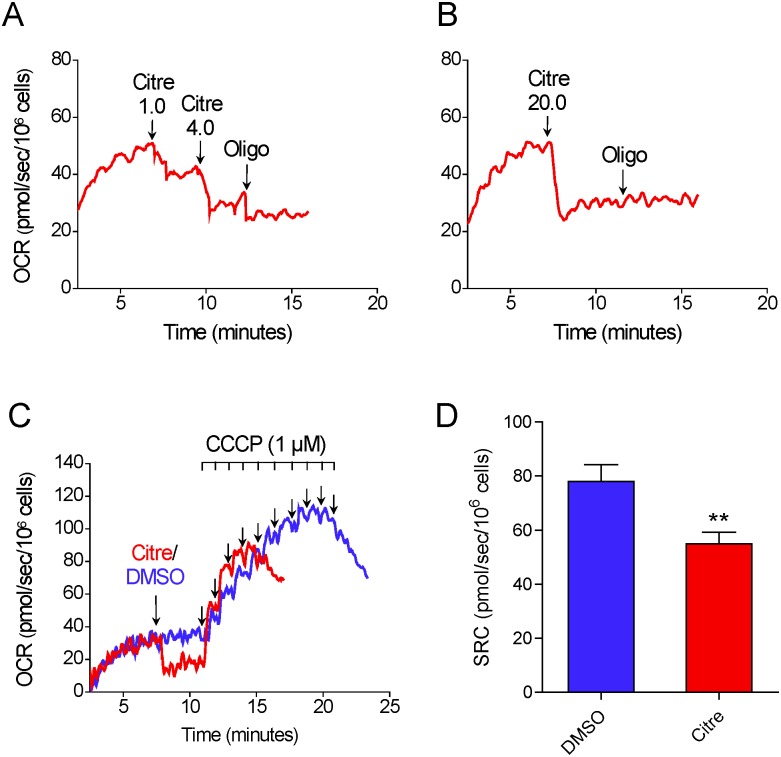
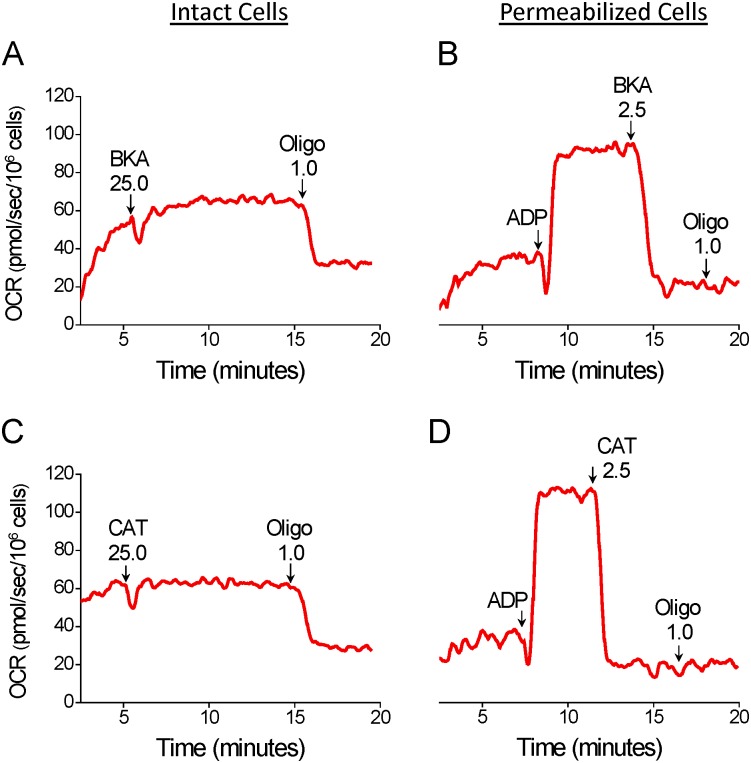
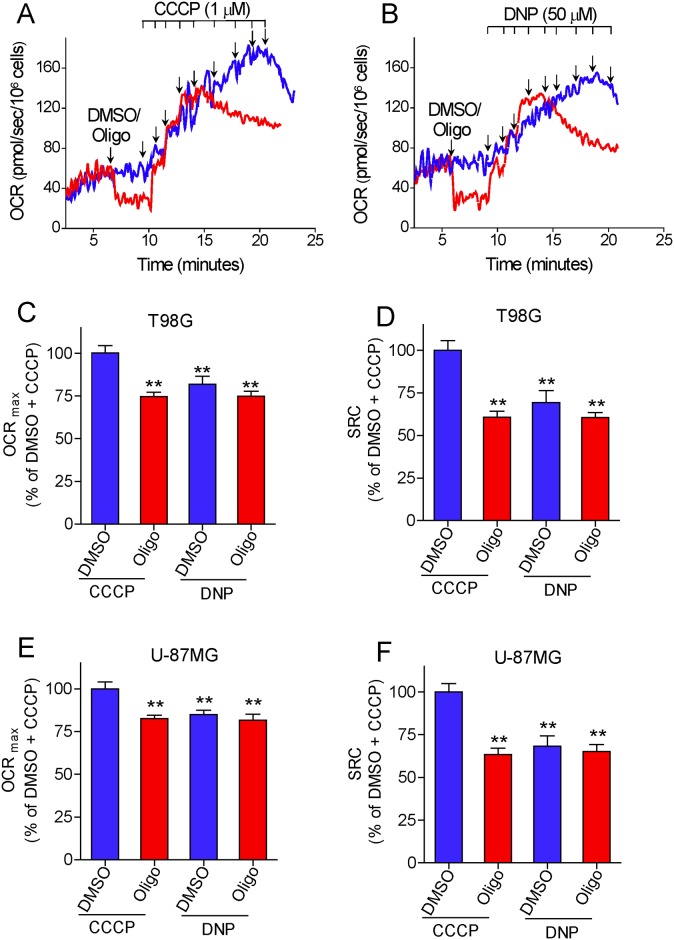
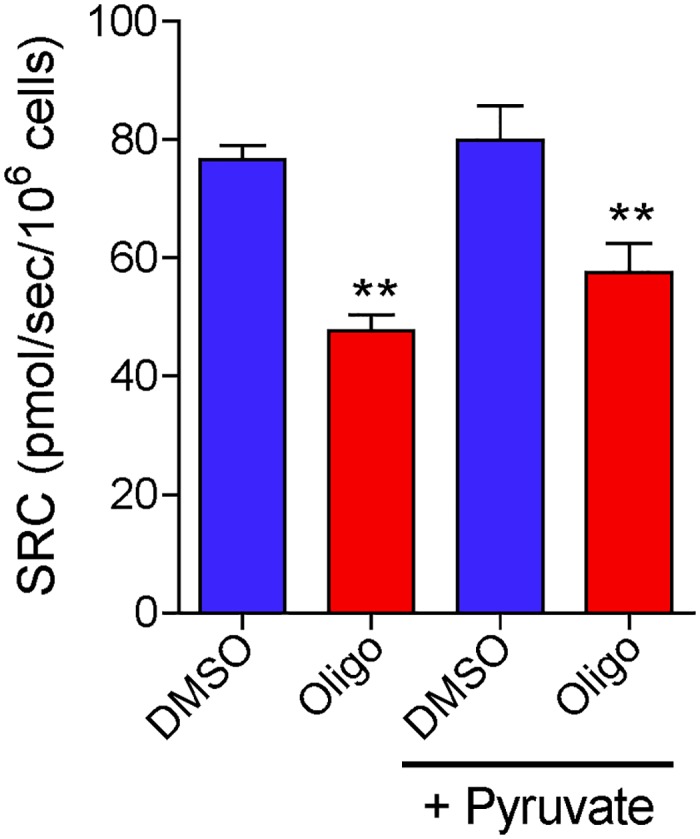
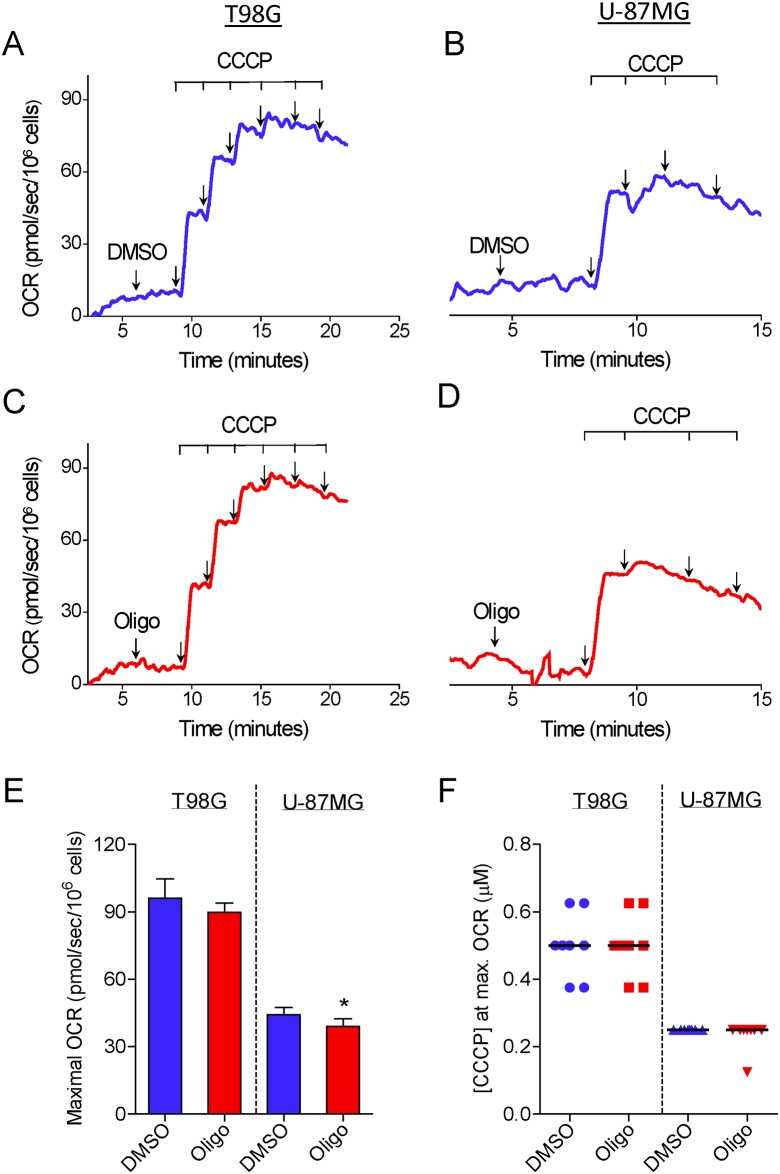
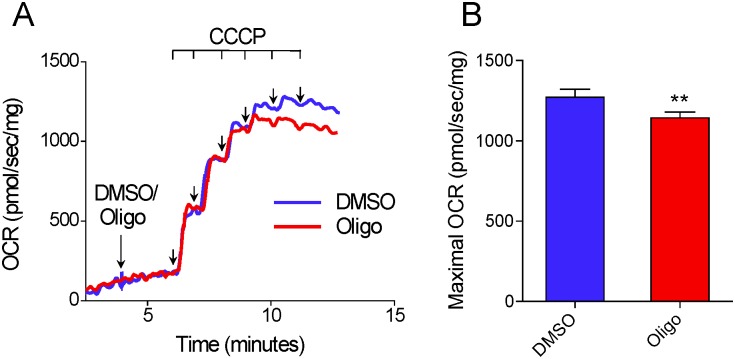
The maximal capacity of the mitochondrial electron transport system (ETS) in intact cells is frequently estimated by promoting protonophore-induced maximal oxygen consumption preceded by inhibition of oxidative phosphorylation by oligomycin. In the present study, human glioma (T98G and U-87MG) and prostate cancer (PC-3) cells were titrated with different concentrations of the protonophore CCCP to induce maximal oxygen consumption rate (OCR) within respirometers in a conventional growth medium. The results demonstrate that the presence of oligomycin or its A-isomer leads to underestimation of maximal ETS capacity. In the presence of oligomycin, the spare respiratory capacity (SRC), i.e., the difference between the maximal and basal cellular OCR, was underestimated by 25 to 45%. The inhibitory effect of oligomycin on SRC was more pronounced in T98G cells and was observed in both suspended and attached cells. Underestimation of SRC also occurred when oxidative phosphorylation was fully inhibited by the ATP synthase inhibitor citreoviridin. Further experiments indicated that oligomycin cannot be replaced by the adenine nucleotide translocase inhibitors bongkrekic acid or carboxyatractyloside because, although these compounds have effects in permeabilized cells, they do not inhibit oxidative phosphorylation in intact cells. We replaced CCCP by FCCP, another potent protonophore and similar results were observed. Lower maximal OCR and SRC values were obtained with the weaker protonophore 2,4-dinitrophenol, and these parameters were not affected by the presence of oligomycin. In permeabilized cells or isolated brain mitochondria incubated with respiratory substrates, only a minor inhibitory effect of oligomycin on CCCP-induced maximal OCR was observed. We conclude that unless a previously validated protocol is employed, maximal ETS capacity in intact cells should be estimated without oligomycin. The inhibitory effect of an ATP synthase blocker on potent protonophore-induced maximal OCR may be associated with impaired metabolism of mitochondrial respiratory substrates.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
