

Heat Induction of Cyclic Electron Flow around Photosystem I in the Symbiotic Dinoflagellate Symbiodinium
- PMID: 26951432
- PMCID: PMC4854689
- DOI: 10.1104/pp.15.01886
Heat Induction of Cyclic Electron Flow around Photosystem I in the Symbiotic Dinoflagellate Symbiodinium
Abstract
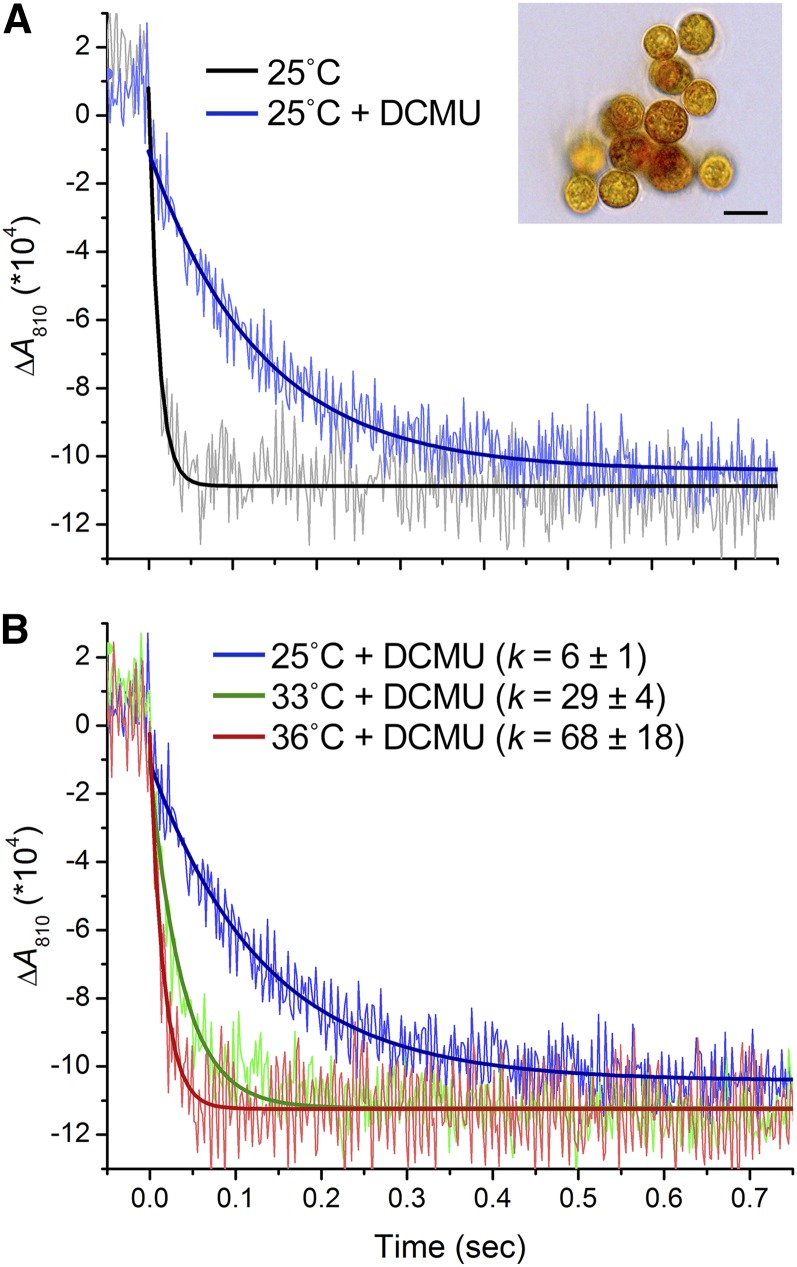
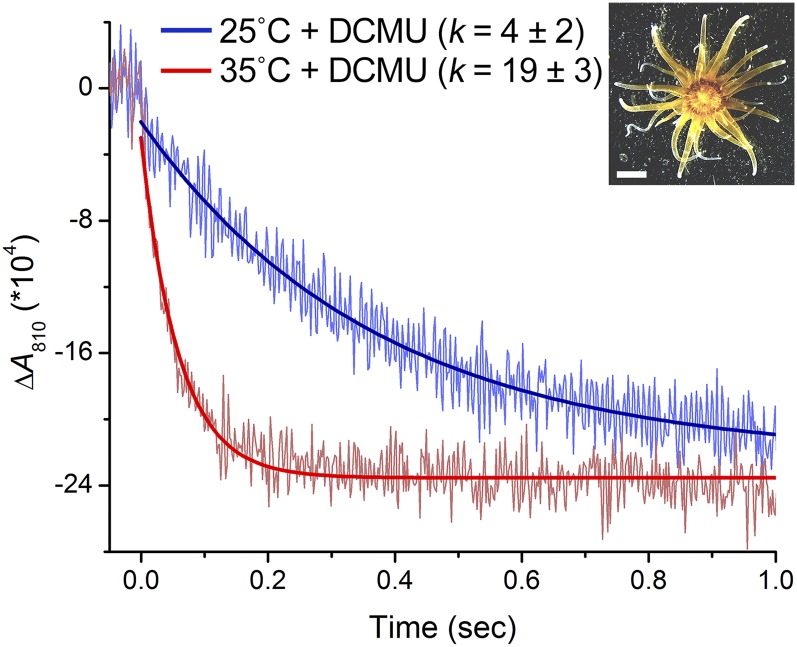
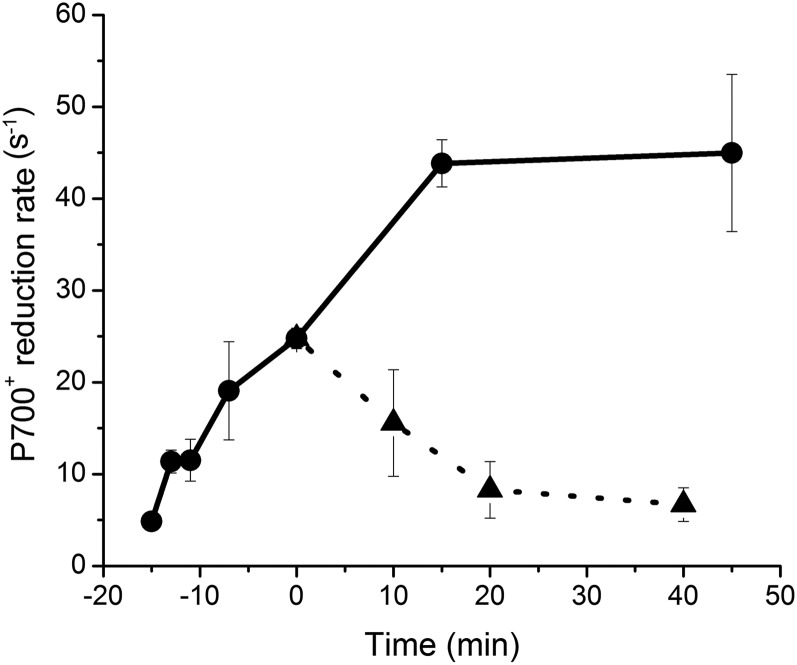
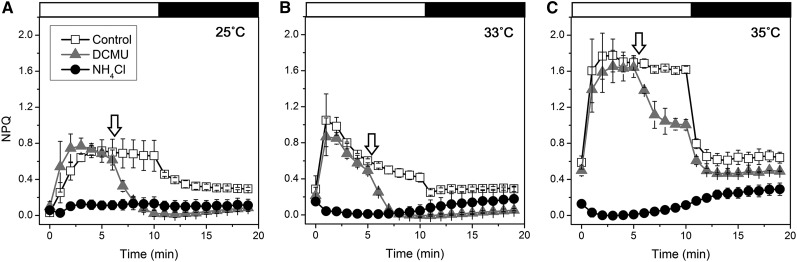
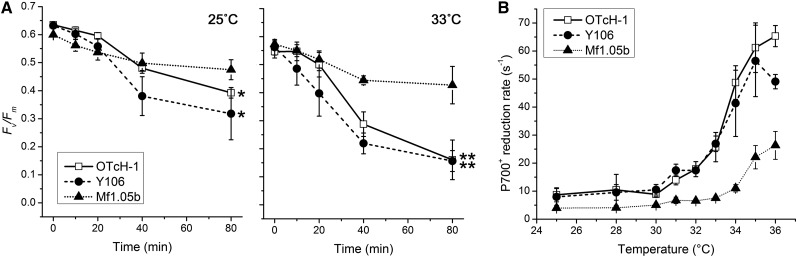
Increases in seawater temperature impair photosynthesis (photoinhibition) in the symbiotic dinoflagellate Symbiodinium within cnidarian hosts, such as corals and sea anemones, and may destroy their symbiotic relationship. Although the degree of photoinhibition in Symbiodinium under heat stress differs among strains, the differences in their responses to increased temperatures, including cyclic electron flow (CEF), which sustains photoprotective thermal energy dissipation, have not been investigated. Here, we examined CEF in cultured Symbiodinium cells or those in an endosymbiotic relationship within a cnidarian host. The light-dependent reduction of the primary electron donor photosystem I, i.e. P700(+), was enhanced in any Symbiodinium cell by increasing temperatures, indicating CEF was induced by heat, which was accompanied by thermal energy dissipation activation. The critical temperatures for inducing CEF were different among Symbiodinium strains. The clade A strains with greater susceptibility to photoinhibition, OTcH-1 and Y106, exhibited higher CEF activities under moderate heat stress than a more phototolerant clade B strain Mf1.05b, suggesting that the observed CEF induction was not a preventive measure but a stress response in Symbiodinium.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Aro EM, Virgin I, Andersson B (1993) Photoinhibition of Photosystem II. Inactivation, protein damage and turnover. Biochim Biophys Acta 1143: 113–134 - PubMed
-
- Breyton C, Nandha B, Johnson GN, Joliot P, Finazzi G (2006) Redox modulation of cyclic electron flow around photosystem I in C3 plants. Biochemistry 45: 13465–13475 - PubMed
-
- Bukhov N, Carpentier R (2004) Alternative photosystem I-driven electron transport routes: mechanisms and functions. Photosynth Res 82: 17–33 - PubMed
-
- Bukhov N, Carpentier R, Samson G (2001) Heterogeneity of Photosystem I reaction centers in barley leaves as related to the donation from stromal reductants. Photosynth Res 70: 273–279 - PubMed
-
- Bukhov NG, Wiese C, Neimanis S, Heber U (1999) Heat sensitivity of chloroplasts and leaves: leakage of protons from thylakoids and reversible activation of cyclic electron transport. Photosynth Res 59: 81–93
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
