

Mechanisms of Selective Autophagy in Normal Physiology and Cancer
- PMID: 26953261
- PMCID: PMC4846542
- DOI: 10.1016/j.jmb.2016.02.027
Mechanisms of Selective Autophagy in Normal Physiology and Cancer
Abstract
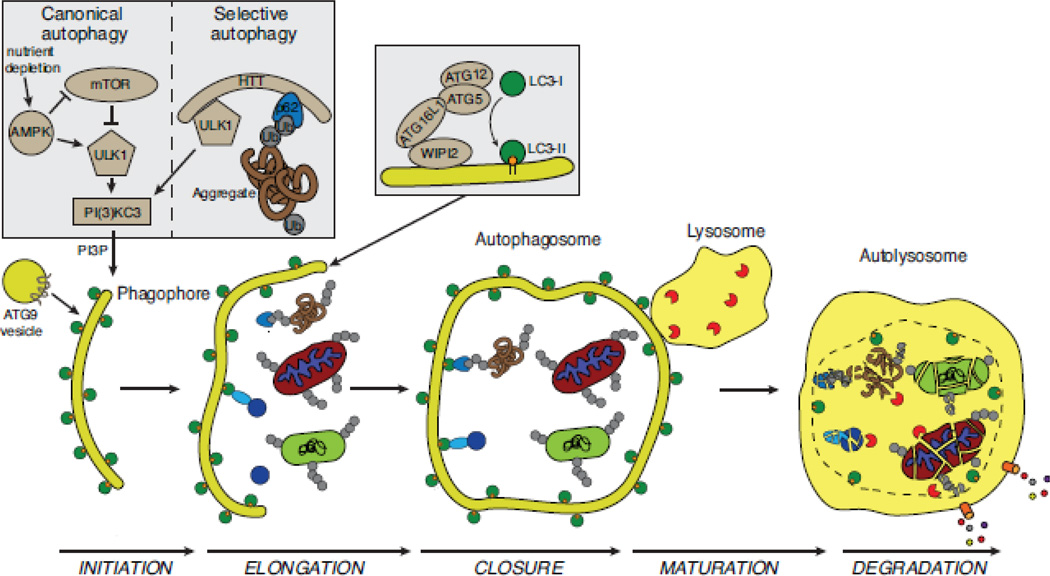
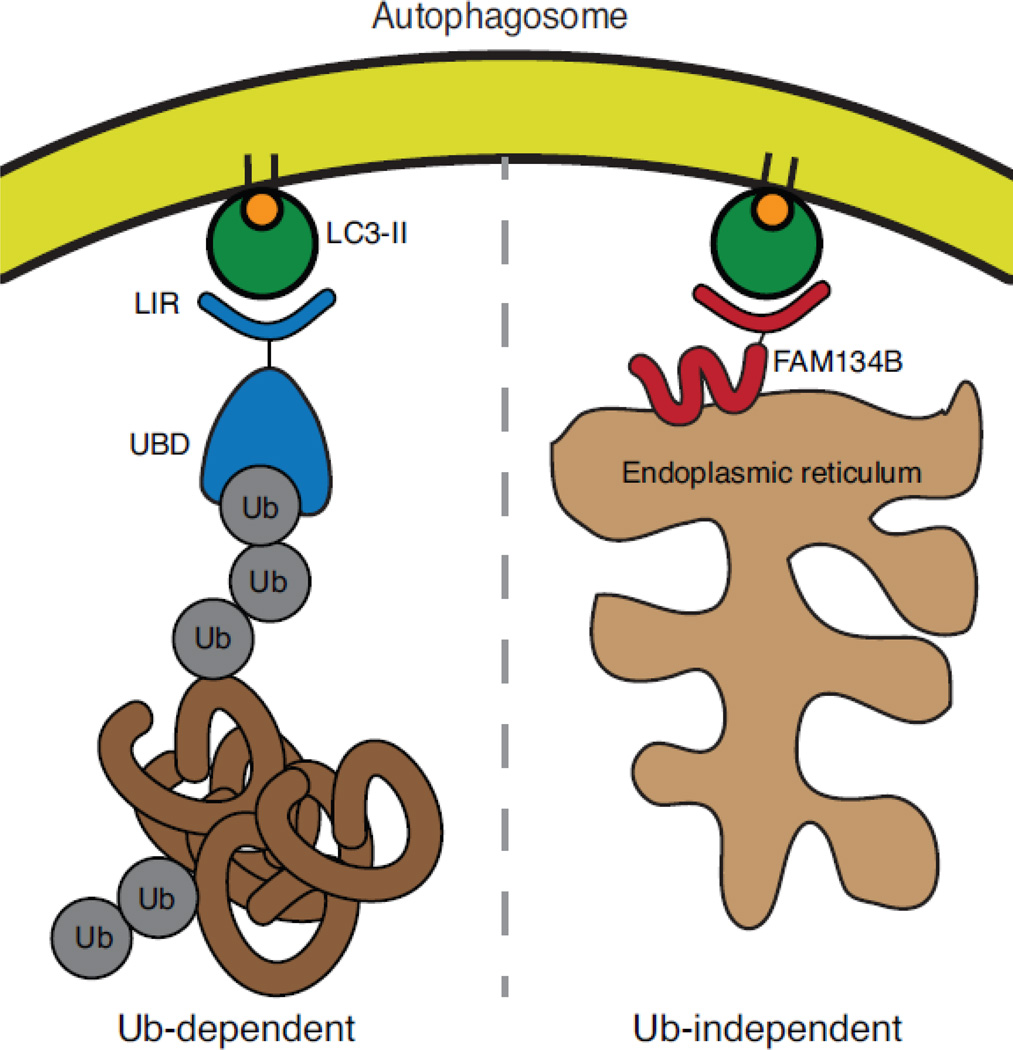
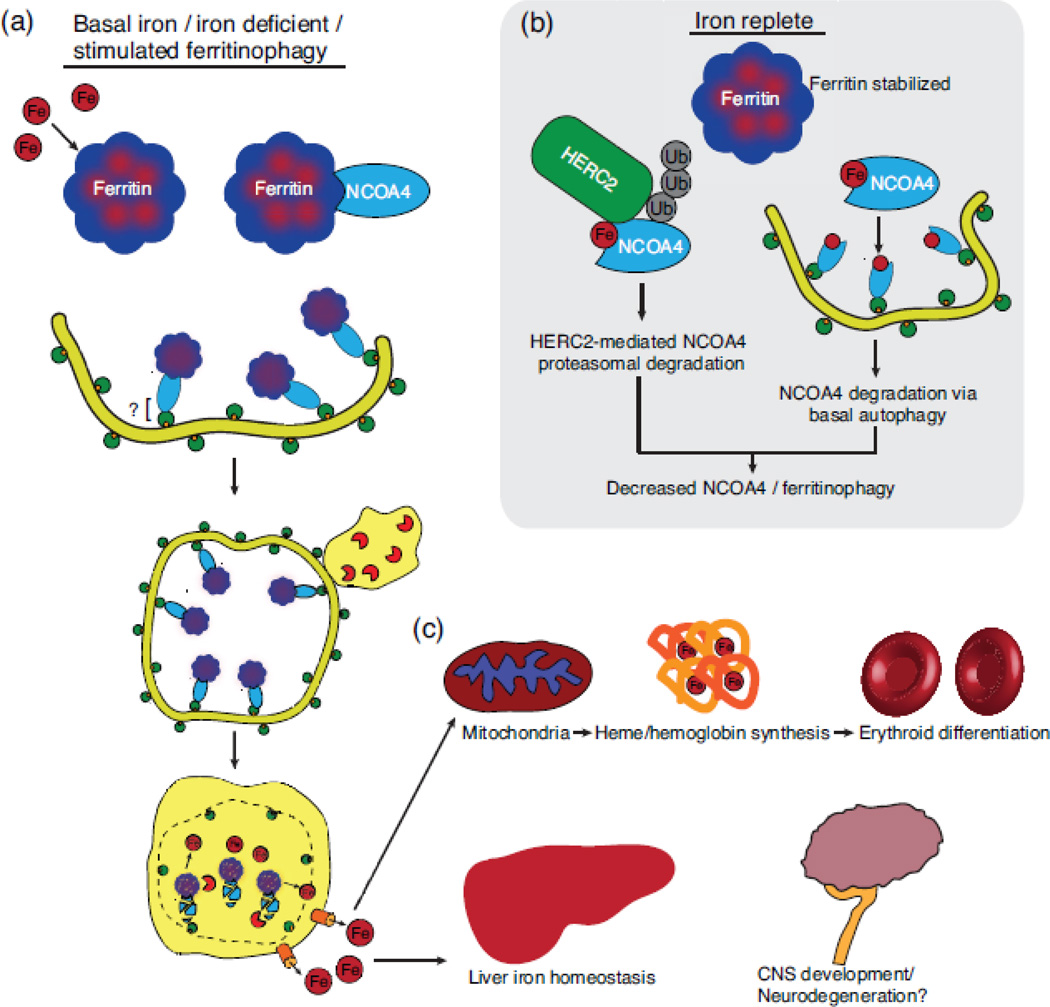
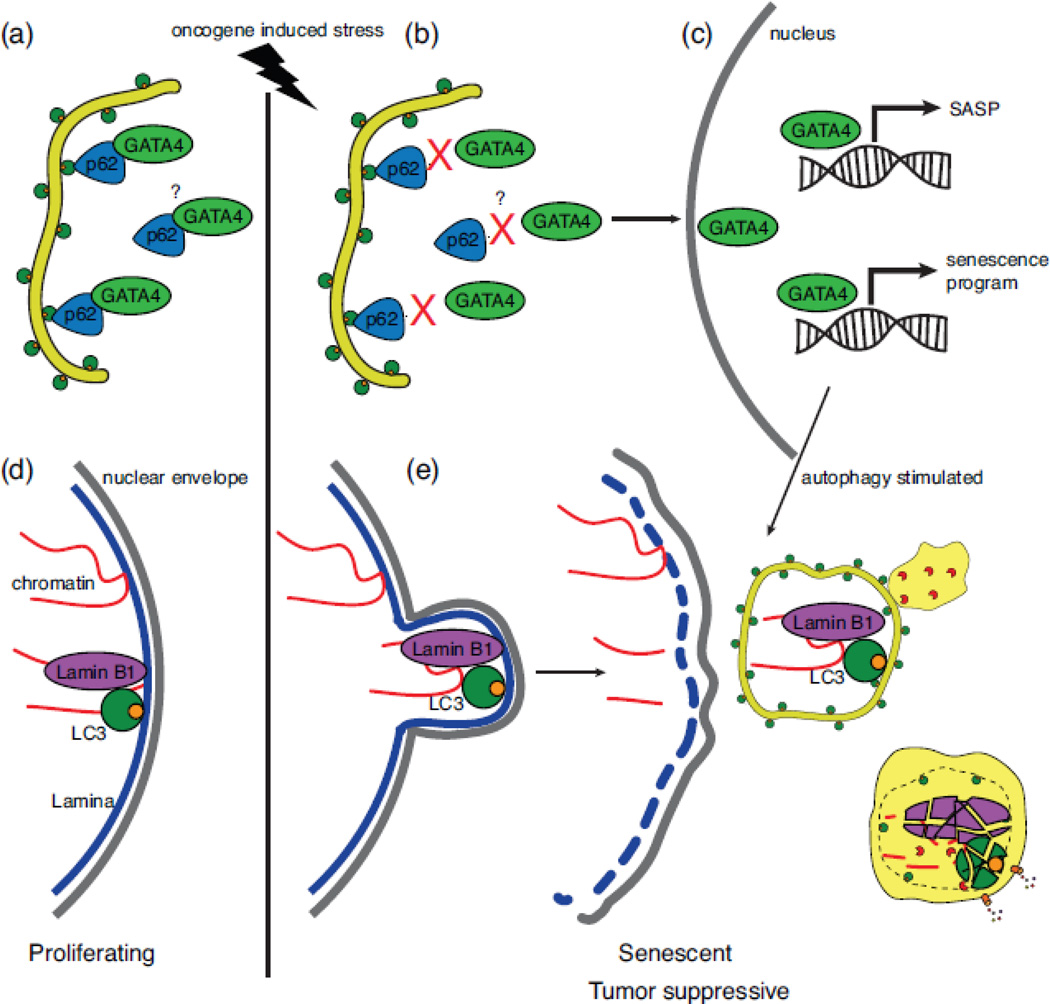
Selective autophagy is critical for regulating cellular homeostasis by mediating lysosomal turnover of a wide variety of substrates including proteins, aggregates, organelles, and pathogens via a growing class of molecules termed selective autophagy receptors. The molecular mechanisms of selective autophagy receptor action and regulation are complex. Selective autophagy receptors link their bound cargo to the autophagosomal membrane by interacting with lipidated ATG8 proteins (LC3/GABARAP) that are intimately associated with the autophagosome membrane. The cargo signals that selective autophagy receptors recognize are diverse but their recognition can be broadly grouped into two classes, ubiquitin-dependent cargo recognition versus ubiquitin-independent. The roles of post-translational modification of selective autophagy receptors in regulating these pathways in response to stimuli are an active area of research. Here we will review recent advances in the identification of selective autophagy receptors and their regulatory mechanisms. Given its importance in maintaining cellular homeostasis, disruption of autophagy can lead to disease including neurodegeneration and cancer. The role of autophagy in cancer is complex as autophagy can mediate promotion or inhibition of tumorigenesis. Here we will also review the importance of autophagy in cancer with a specific focus on the role of selective autophagy receptors.
Keywords: cancer; ferritinophagy; macroautophagy (autophagy); mitophagy; selective autophagy.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Mizushima N, Yoshimori T, Ohsumi Y. The Role of Atg Proteins in Autophagosome Formation. Annu. Rev. Cell Dev. Biol. 2011;27:107–132. - PubMed
-
- Noda NN, Inagaki F. Mechanisms of Autophagy. Annu. Rev. Biophys. 2015;44:101–122. - PubMed
-
- Svenning S, Johansen T. Selective autophagy. Essays Biochem. 2013;55:79–92. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
