

Regulatory (pan-)genome of an obligate intracellular pathogen in the PVC superphylum
- PMID: 26953603
- PMCID: PMC4989314
- DOI: 10.1038/ismej.2016.23
Regulatory (pan-)genome of an obligate intracellular pathogen in the PVC superphylum
Abstract
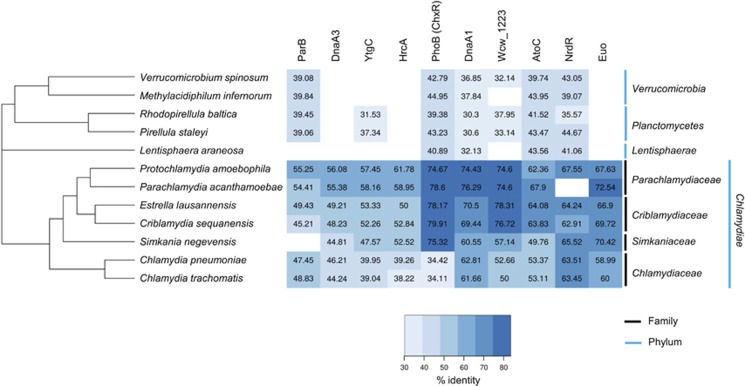
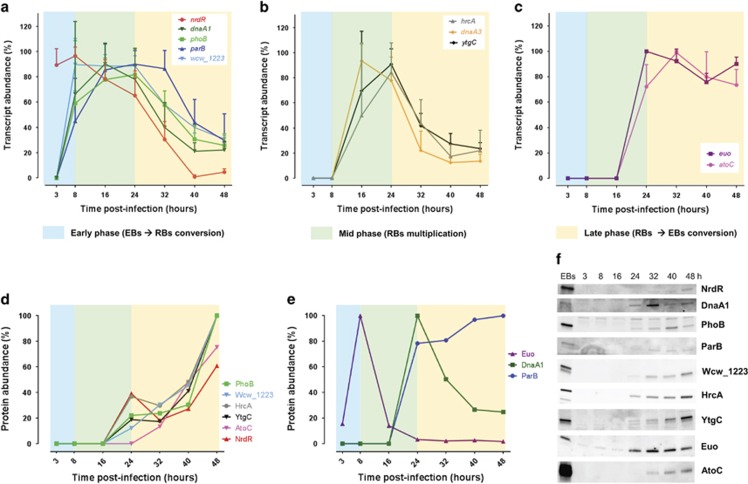
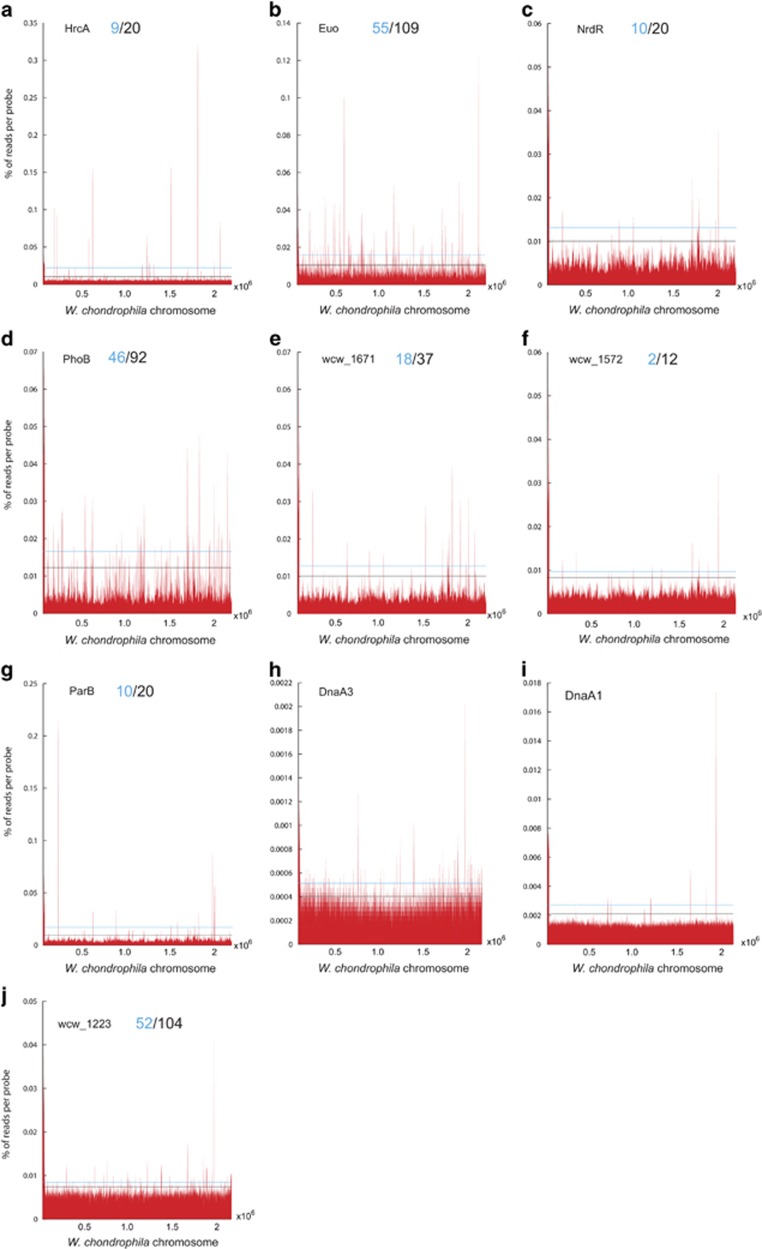
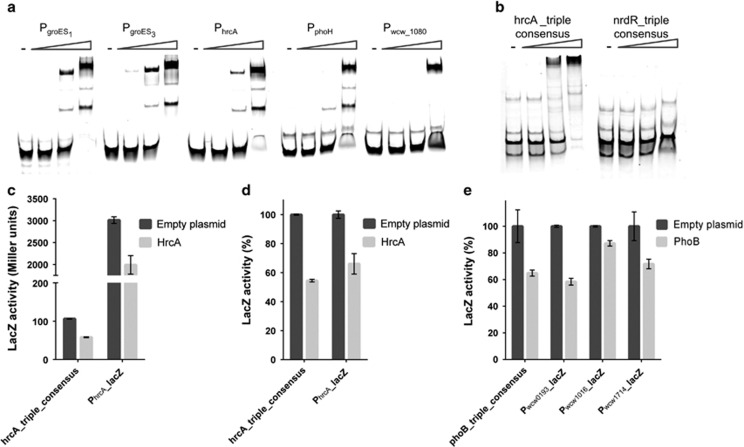
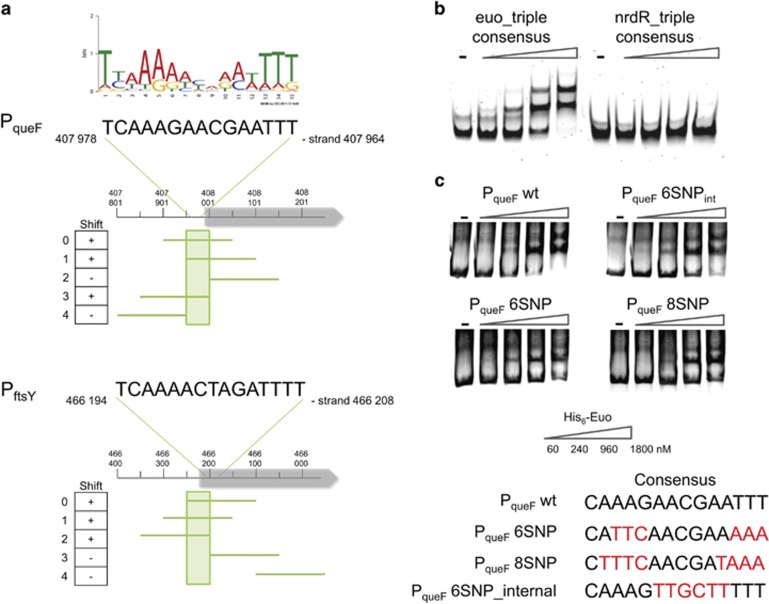
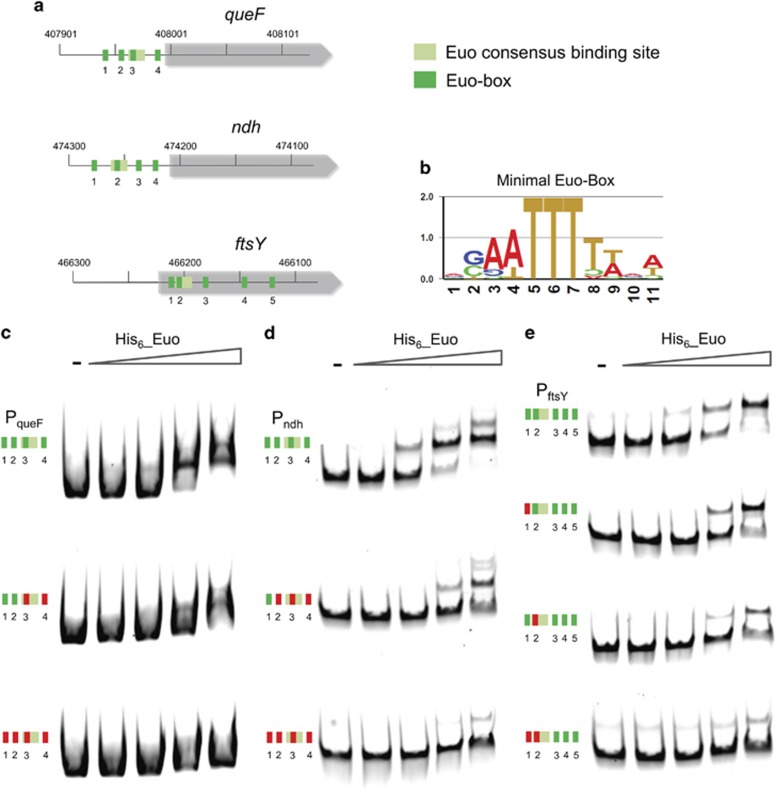
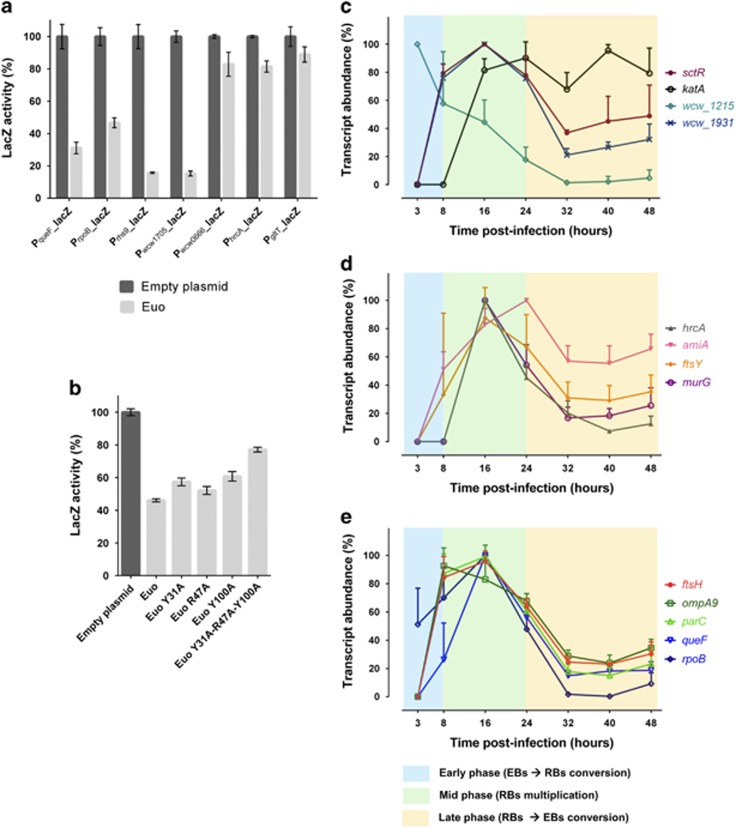
Like other obligate intracellular bacteria, the Chlamydiae feature a compact regulatory genome that remains uncharted owing to poor genetic tractability. Exploiting the reduced number of transcription factors (TFs) encoded in the chlamydial (pan-)genome as a model for TF control supporting the intracellular lifestyle, we determined the conserved landscape of TF specificities by ChIP-Seq (chromatin immunoprecipitation-sequencing) in the chlamydial pathogen Waddlia chondrophila. Among 10 conserved TFs, Euo emerged as a master TF targeting >100 promoters through conserved residues in a DNA excisionase-like winged helix-turn-helix-like (wHTH) fold. Minimal target (Euo) boxes were found in conserved developmentally-regulated genes governing vertical genome transmission (cytokinesis and DNA replication) and genome plasticity (transposases). Our ChIP-Seq analysis with intracellular bacteria not only reveals that global TF regulation is maintained in the reduced regulatory genomes of Chlamydiae, but also predicts that master TFs interpret genomic information in the obligate intracellular α-proteobacteria, including the rickettsiae, from which modern day mitochondria evolved.
Figures
Similar articles
-
Identification of new DNA-associated proteins from Waddlia chondrophila.Sci Rep. 2019 Mar 20;9(1):4885. doi: 10.1038/s41598-019-40732-1. Sci Rep. 2019. PMID: 30894592 Free PMC article.
-
Unity in variety--the pan-genome of the Chlamydiae.Mol Biol Evol. 2011 Dec;28(12):3253-70. doi: 10.1093/molbev/msr161. Epub 2011 Jun 20. Mol Biol Evol. 2011. PMID: 21690563 Free PMC article.
-
Proteins Related to the Type I Secretion System Are Associated with Secondary SecA_DEAD Domain Proteins in Some Species of Planctomycetes, Verrucomicrobia, Proteobacteria, Nitrospirae and Chlorobi.PLoS One. 2015 Jun 1;10(6):e0129066. doi: 10.1371/journal.pone.0129066. eCollection 2015. PLoS One. 2015. PMID: 26030905 Free PMC article.
-
Insight in the biology of Chlamydia-related bacteria.Microbes Infect. 2018 Aug-Sep;20(7-8):432-440. doi: 10.1016/j.micinf.2017.11.008. Epub 2017 Dec 18. Microbes Infect. 2018. PMID: 29269129 Review.
-
The PVC superphylum: exceptions to the bacterial definition?Antonie Van Leeuwenhoek. 2013 Oct;104(4):451-66. doi: 10.1007/s10482-013-9986-1. Epub 2013 Aug 4. Antonie Van Leeuwenhoek. 2013. PMID: 23912444 Review.
Cited by
-
A SpoIID Homolog Cleaves Glycan Strands at the Chlamydial Division Septum.mBio. 2019 Jul 16;10(4):e01128-19. doi: 10.1128/mBio.01128-19. mBio. 2019. PMID: 31311880 Free PMC article.
-
Genomewide Transcriptional Responses of Iron-Starved Chlamydia trachomatis Reveal Prioritization of Metabolic Precursor Synthesis over Protein Translation.mSystems. 2018 Feb 13;3(1):e00184-17. doi: 10.1128/mSystems.00184-17. eCollection 2018 Jan-Feb. mSystems. 2018. PMID: 29468197 Free PMC article.
-
Environmental Metagenomic Assemblies Reveal Seven New Highly Divergent Chlamydial Lineages and Hallmarks of a Conserved Intracellular Lifestyle.Front Microbiol. 2018 Feb 20;9:79. doi: 10.3389/fmicb.2018.00079. eCollection 2018. Front Microbiol. 2018. PMID: 29515524 Free PMC article.
-
Trends in Symbiont-Induced Host Cellular Differentiation.Results Probl Cell Differ. 2020;69:137-176. doi: 10.1007/978-3-030-51849-3_5. Results Probl Cell Differ. 2020. PMID: 33263871 Free PMC article.
-
Robust Heat Shock Response in Chlamydia Lacking a Typical Heat Shock Sigma Factor.Front Microbiol. 2022 Jan 3;12:812448. doi: 10.3389/fmicb.2021.812448. eCollection 2021. Front Microbiol. 2022. PMID: 35046926 Free PMC article.
References
-
- Baud D, Regan L, Greub G. (2008). Emerging role of Chlamydia and Chlamydia-like organisms in adverse pregnancy outcomes. Curr Opin Infect Dis 21: 70–76. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
