

Amino Acids Rather than Glucose Account for the Majority of Cell Mass in Proliferating Mammalian Cells
- PMID: 26954548
- PMCID: PMC4766004
- DOI: 10.1016/j.devcel.2016.02.012
Amino Acids Rather than Glucose Account for the Majority of Cell Mass in Proliferating Mammalian Cells
Abstract
Cells must duplicate their mass in order to proliferate. Glucose and glutamine are the major nutrients consumed by proliferating mammalian cells, but the extent to which these and other nutrients contribute to cell mass is unknown. We quantified the fraction of cell mass derived from different nutrients and found that the majority of carbon mass in cells is derived from other amino acids, which are consumed at much lower rates than glucose and glutamine. While glucose carbon has diverse fates, glutamine contributes most to protein, suggesting that glutamine's ability to replenish tricarboxylic acid cycle intermediates (anaplerosis) is primarily used for amino acid biosynthesis. These findings demonstrate that rates of nutrient consumption are indirectly associated with mass accumulation and suggest that high rates of glucose and glutamine consumption support rapid cell proliferation beyond providing carbon for biosynthesis.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
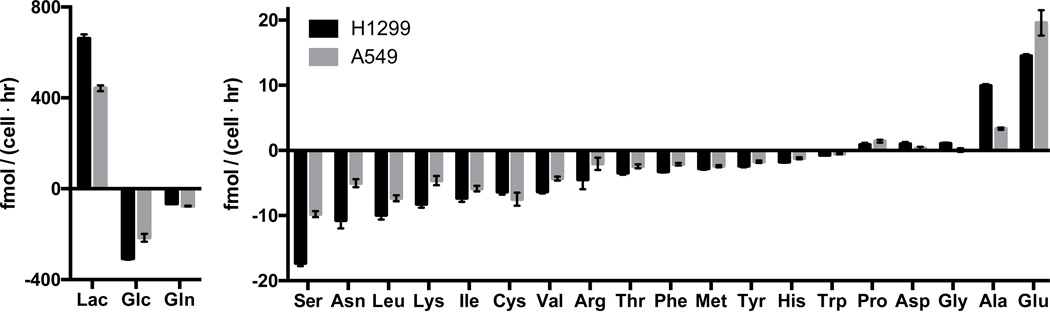
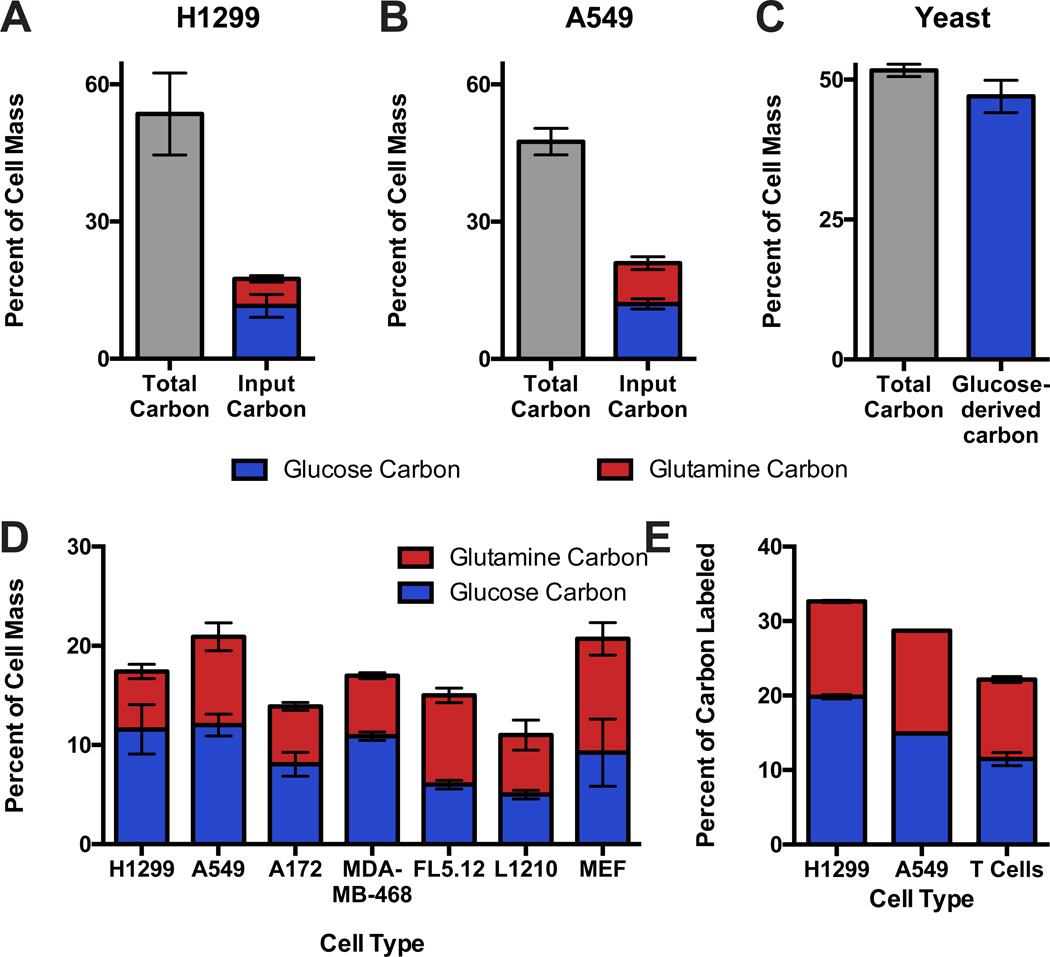
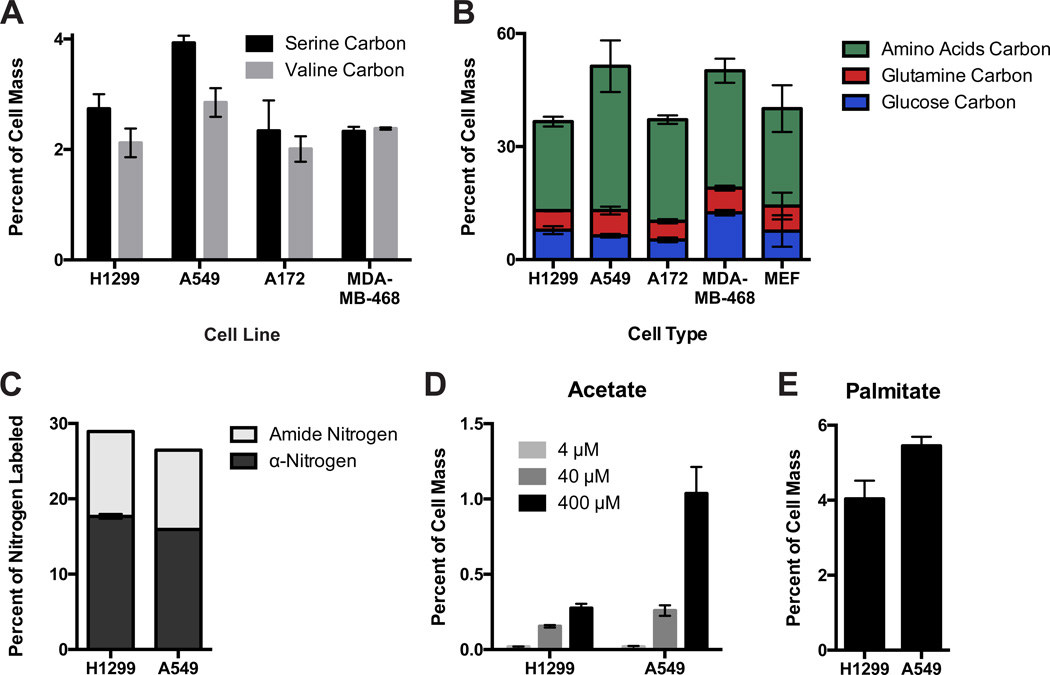
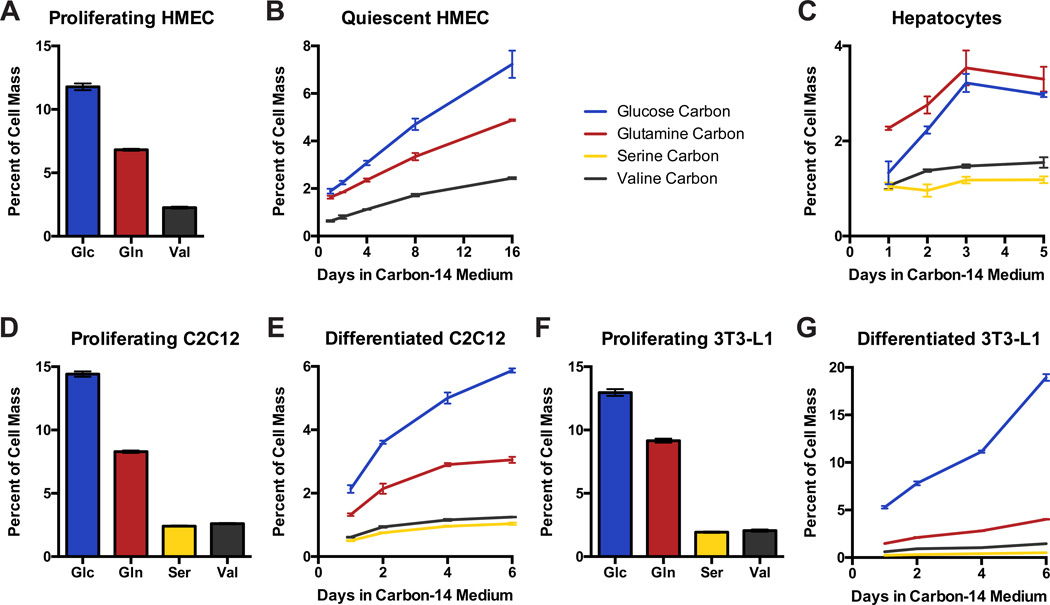
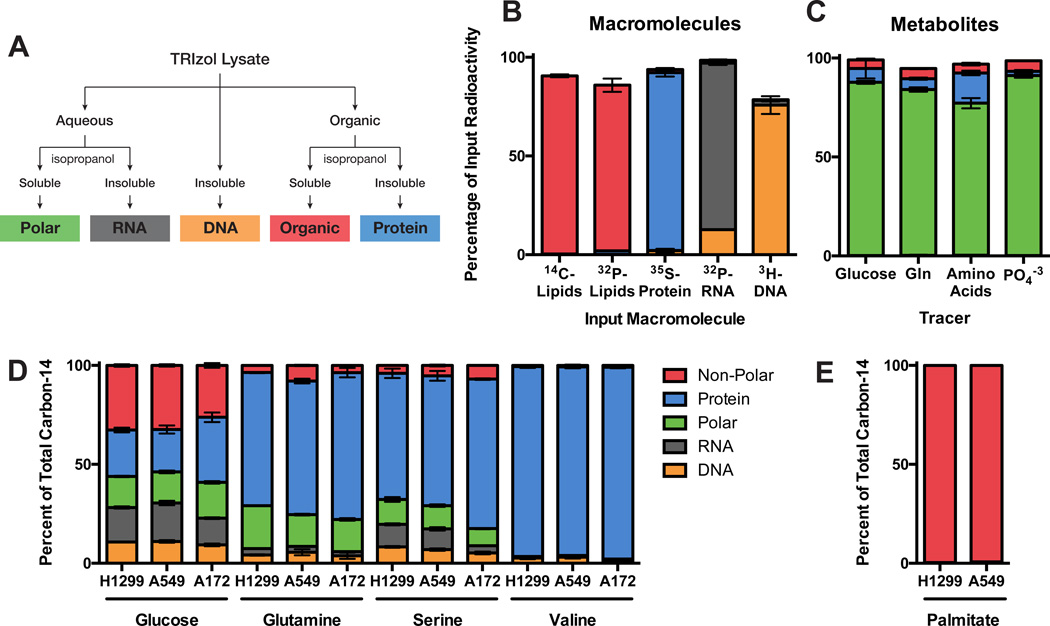
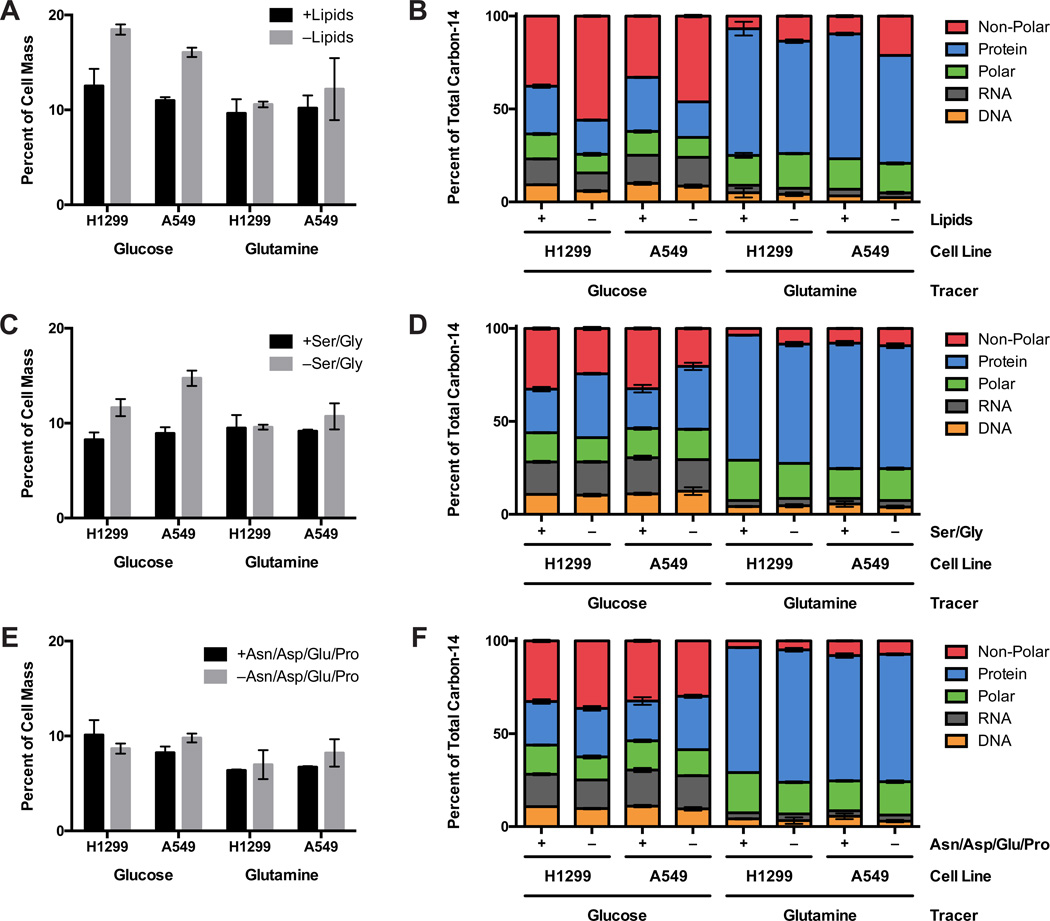
Comment in
-
You Are What You Eat... or Are You?Dev Cell. 2016 Mar 7;36(5):483-5. doi: 10.1016/j.devcel.2016.02.021. Dev Cell. 2016. PMID: 26954542
References
-
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the Cell. 5. New York, NY: Garland Science; 2008.
-
- Bailey JM, Howard BV, Dunbar LM, Tillman SF. Control of lipid metabolism in cultured cells. Lipids. 1972;7:125–134. - PubMed
-
- Bonarius HP, Hatzimanikatis V, Meesters KP, de Gooijer CD, Schmid G, Tramper J. Metabolic flux analysis of hybridoma cells in different culture media using mass balances. Biotechnology and bioengineering. 1996;50:299–318. - PubMed
-
- Boulter D, Barber JT. Amino-acid metabolism in germinating seeds of Vicia faba L. in relation to their biology. New Phytologist. 1963;62:301–316.
Publication types
MeSH terms
Substances
Grants and funding
- K08DK090147/DK/NIDDK NIH HHS/United States
- R21 CA198028/CA/NCI NIH HHS/United States
- P30CA14051/CA/NCI NIH HHS/United States
- R01 HL108006/HL/NHLBI NIH HHS/United States
- U54CA121852/CA/NCI NIH HHS/United States
- R01 CA168653/CA/NCI NIH HHS/United States
- R01HL108006/HL/NHLBI NIH HHS/United States
- P30 CA014051/CA/NCI NIH HHS/United States
- R21 CA194829/CA/NCI NIH HHS/United States
- R01 DK105550/DK/NIDDK NIH HHS/United States
- R01CA168653/CA/NCI NIH HHS/United States
- U54 CA143874/CA/NCI NIH HHS/United States
- U54CA143874/CA/NCI NIH HHS/United States
- U54 CA121852/CA/NCI NIH HHS/United States
- Howard Hughes Medical Institute/United States
- K08 DK090147/DK/NIDDK NIH HHS/United States
- T32 GM007105/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
