

Molecular circuitry of stem cell fate in skeletal muscle regeneration, ageing and disease
- PMID: 26956195
- PMCID: PMC4918817
- DOI: 10.1038/nrm.2016.7
Molecular circuitry of stem cell fate in skeletal muscle regeneration, ageing and disease
Abstract
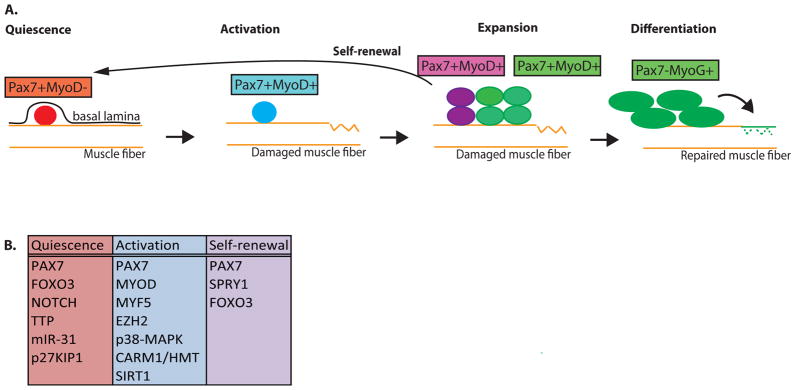
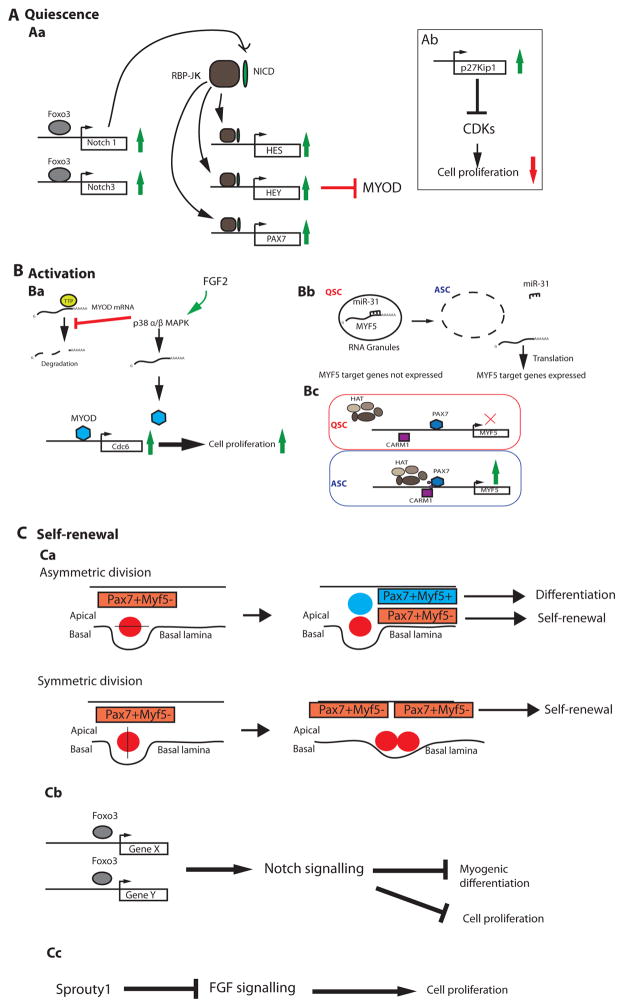
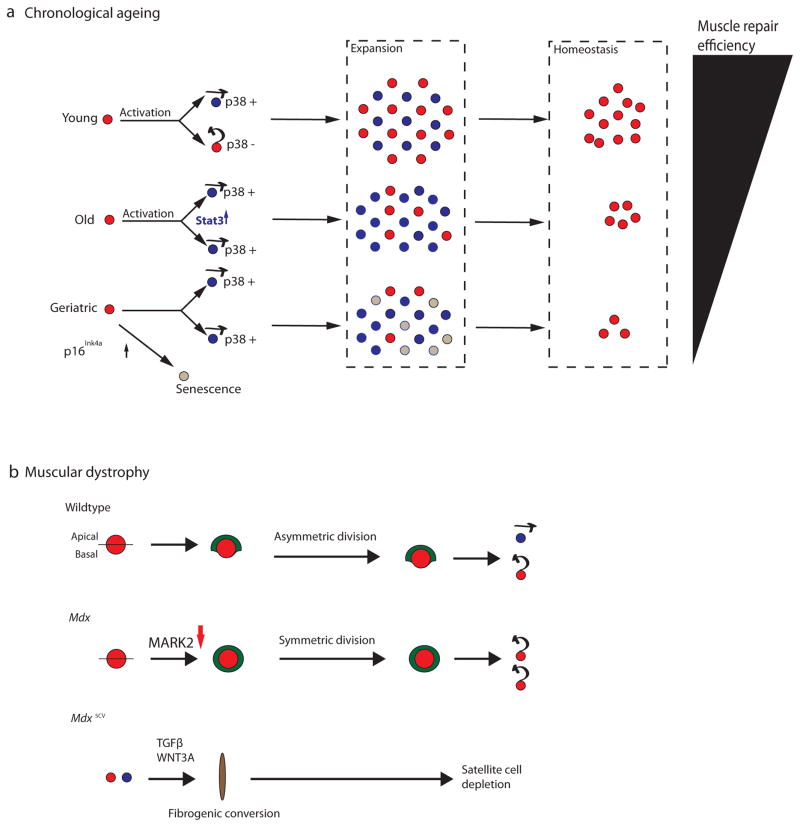
Satellite cells are adult myogenic stem cells that repair damaged muscle. The enduring capacity for muscle regeneration requires efficient satellite cell expansion after injury, their differentiation to produce myoblasts that can reconstitute damaged fibres and their self-renewal to replenish the muscle stem cell pool for subsequent rounds of injury and repair. Emerging studies indicate that misregulation of satellite cell fate and function can contribute to age-associated muscle dysfunction and influence the severity of muscle diseases, including Duchenne muscular dystrophy (DMD). It has also become apparent that satellite cell fate during muscle regeneration and ageing, and in the context of DMD, is governed by an intricate network of intrinsic and extrinsic regulators. Targeted manipulation of this network may offer unique opportunities for muscle regenerative medicine.
Figures
References
-
- Brohl D, et al. Colonization of the satellite cell niche by skeletal muscle progenitor cells depends on Notch signals. Dev Cell. 2012;23:469–81. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
