

MreB-Dependent Organization of the E. coli Cytoplasmic Membrane Controls Membrane Protein Diffusion
- PMID: 26958890
- PMCID: PMC4788719
- DOI: 10.1016/j.bpj.2016.01.010
MreB-Dependent Organization of the E. coli Cytoplasmic Membrane Controls Membrane Protein Diffusion
Abstract
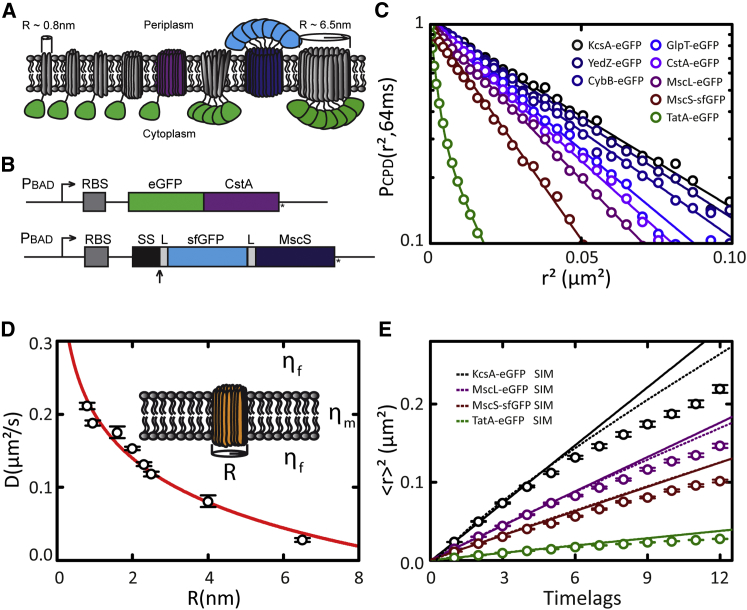
The functional organization of prokaryotic cell membranes, which is essential for many cellular processes, has been challenging to analyze due to the small size and nonflat geometry of bacterial cells. Here, we use single-molecule fluorescence microscopy and three-dimensional quantitative analyses in live Escherichia coli to demonstrate that its cytoplasmic membrane contains microdomains with distinct physical properties. We show that the stability of these microdomains depends on the integrity of the MreB cytoskeletal network underneath the membrane. We explore how the interplay between cytoskeleton and membrane affects trans-membrane protein (TMP) diffusion and reveal that the mobility of the TMPs tested is subdiffusive, most likely caused by confinement of TMP mobility by the submembranous MreB network. Our findings demonstrate that the dynamic architecture of prokaryotic cell membranes is controlled by the MreB cytoskeleton and regulates the mobility of TMPs.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
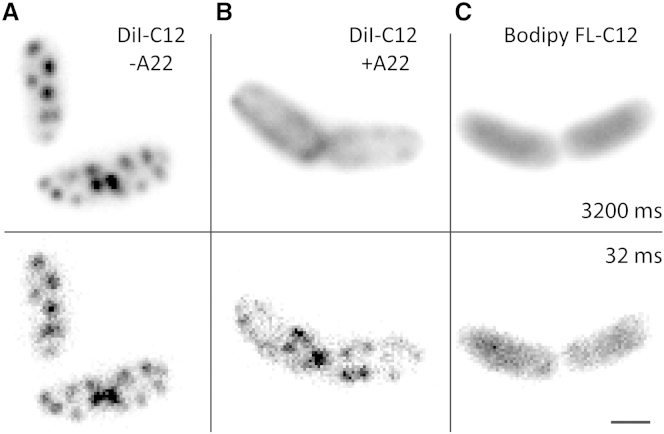
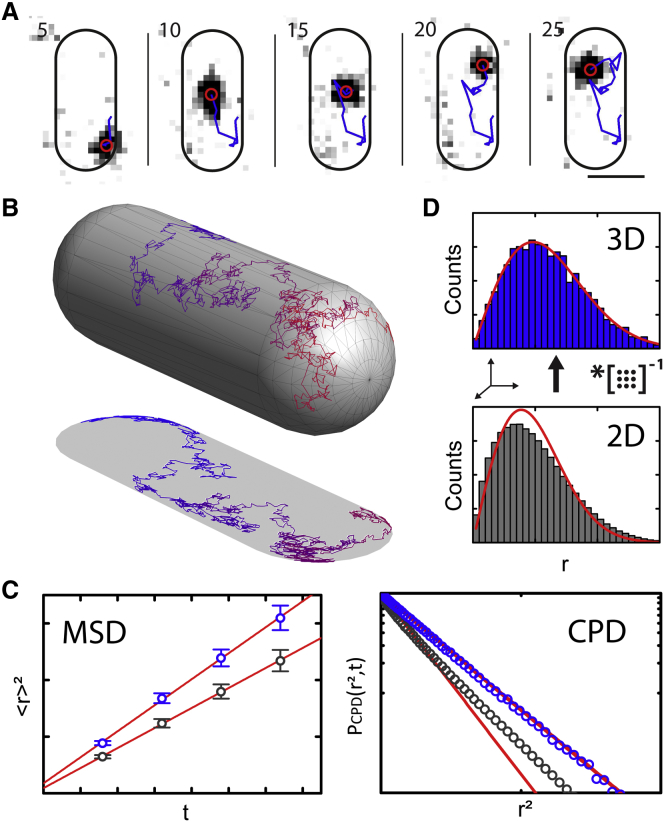
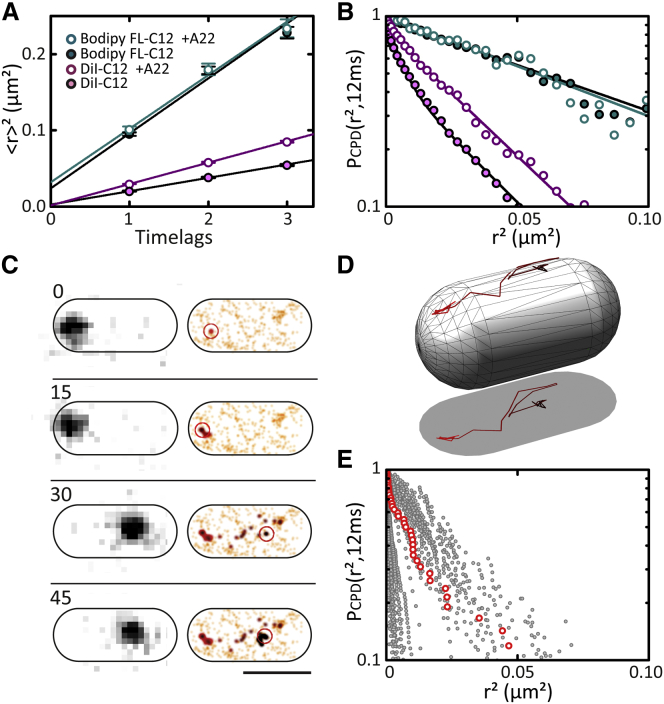
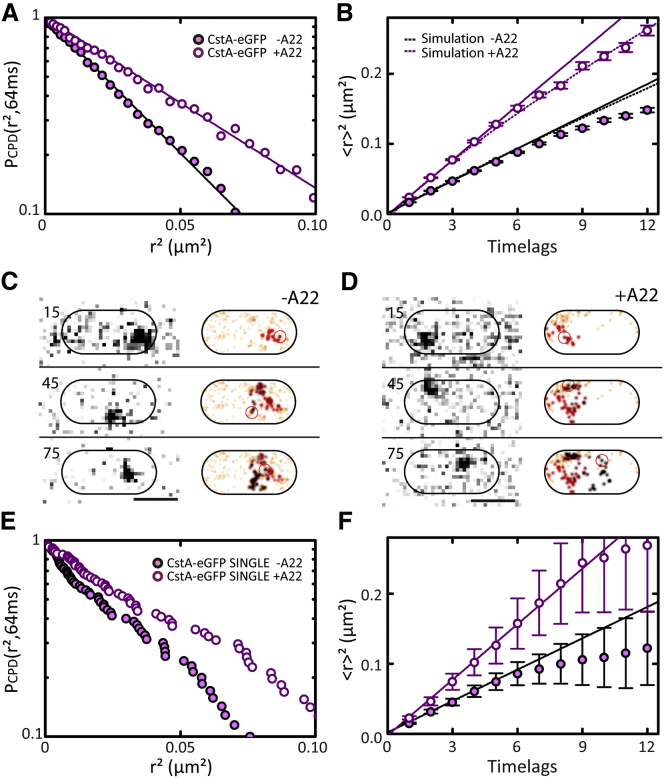
Comment in
-
Spatiotemporal Control of Transmembrane Proteins through the Cytoskeleton: An Evolving Story.Biophys J. 2016 Mar 8;110(5):1036-7. doi: 10.1016/j.bpj.2016.01.011. Biophys J. 2016. PMID: 26958880 Free PMC article. No abstract available.
References
-
- Errington J. Bacterial morphogenesis and the enigmatic MreB helix. Nat. Rev. Microbiol. 2015;13:241–248. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
