

NMR Methods to Study Dynamic Allostery
- PMID: 26964042
- PMCID: PMC4786136
- DOI: 10.1371/journal.pcbi.1004620
NMR Methods to Study Dynamic Allostery
Abstract
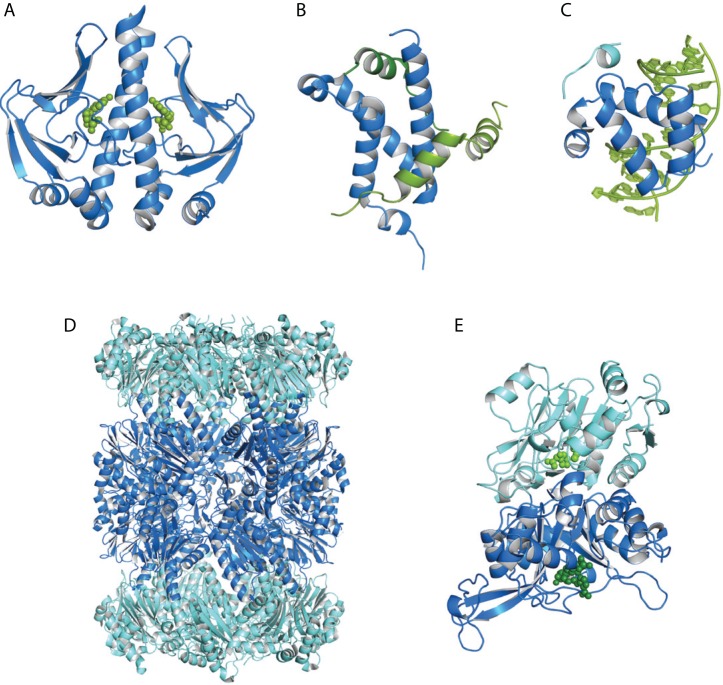
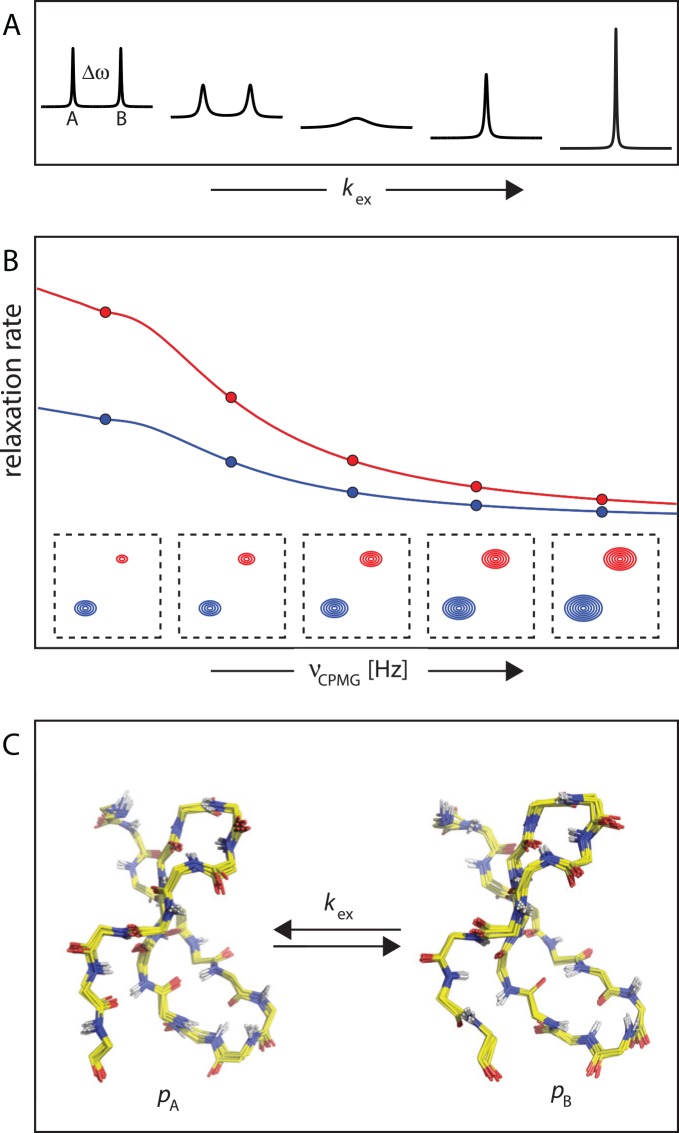
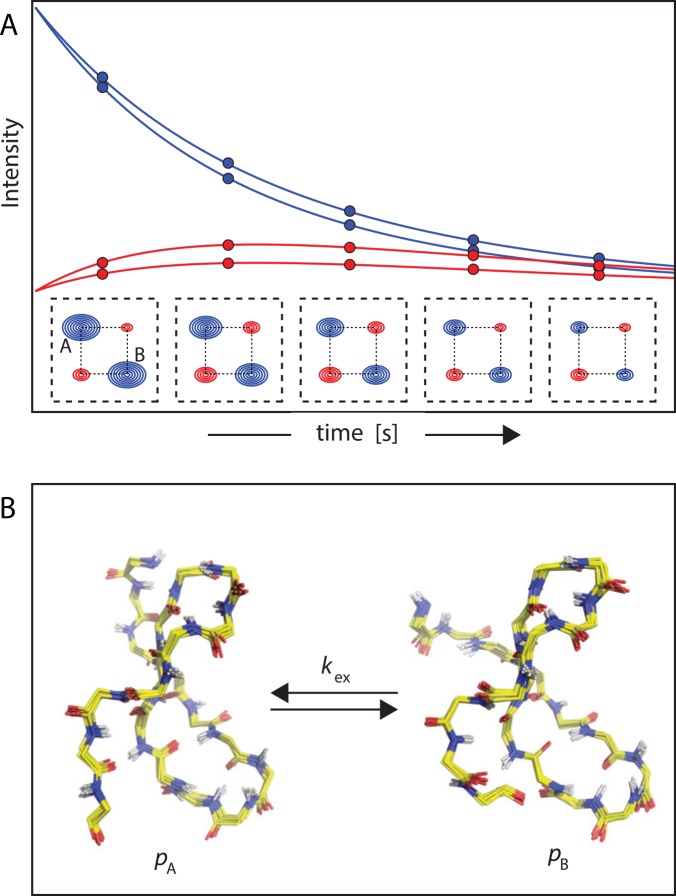
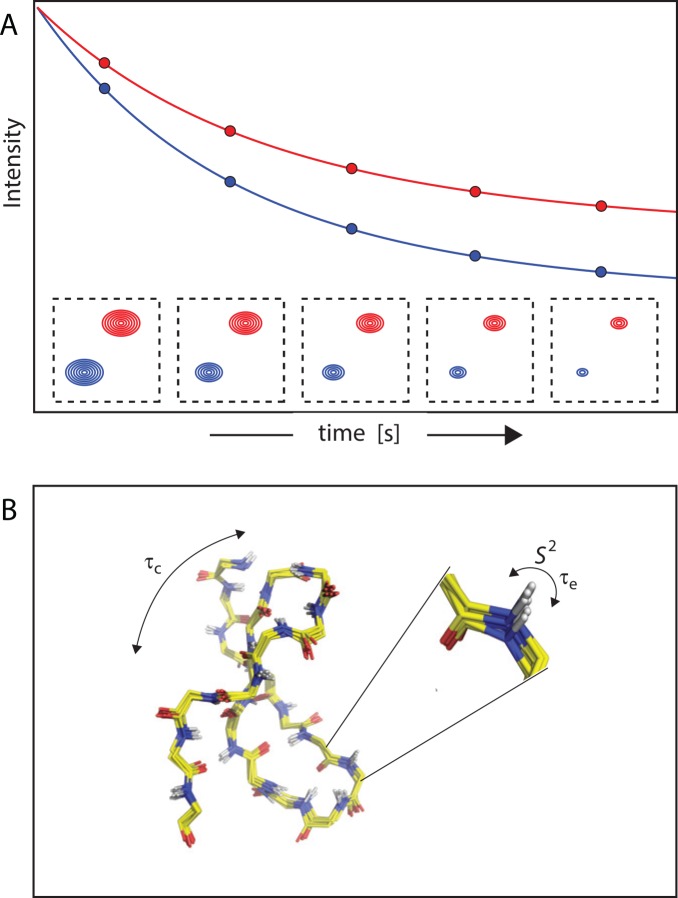
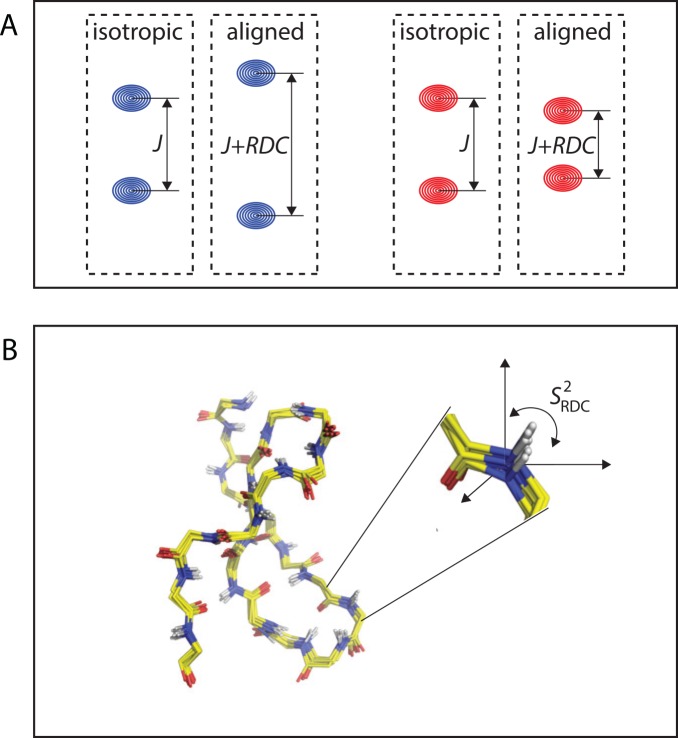
Nuclear magnetic resonance (NMR) spectroscopy provides a unique toolbox of experimental probes for studying dynamic processes on a wide range of timescales, ranging from picoseconds to milliseconds and beyond. Along with NMR hardware developments, recent methodological advancements have enabled the characterization of allosteric proteins at unprecedented detail, revealing intriguing aspects of allosteric mechanisms and increasing the proportion of the conformational ensemble that can be observed by experiment. Here, we present an overview of NMR spectroscopic methods for characterizing equilibrium fluctuations in free and bound states of allosteric proteins that have been most influential in the field. By combining NMR experimental approaches with molecular simulations, atomistic-level descriptions of the mechanisms by which allosteric phenomena take place are now within reach.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Gunasekaran K, Ma B, Nussinov R. Is allostery an intrinsic property of all dynamic proteins? Proteins. 2004;57(3):433–43. - PubMed
-
- Kern D, Zuiderweg ER. The role of dynamics in allosteric regulation. Curr Opin Struct Biol. 2003;13(6):748–57. - PubMed
-
- Swain JF, Gierasch LM. The changing landscape of protein allostery. Curr Opin Struct Biol. 2006;16(1):102–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
