

Cytochrome c folds through foldon-dependent native-like intermediates in an ordered pathway
- PMID: 26966231
- PMCID: PMC4833275
- DOI: 10.1073/pnas.1522674113
Cytochrome c folds through foldon-dependent native-like intermediates in an ordered pathway
Abstract
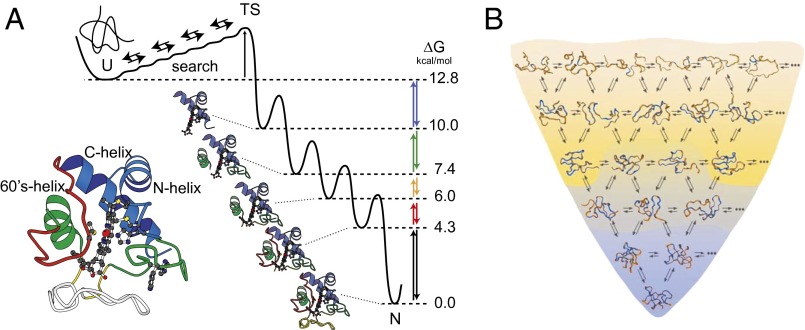
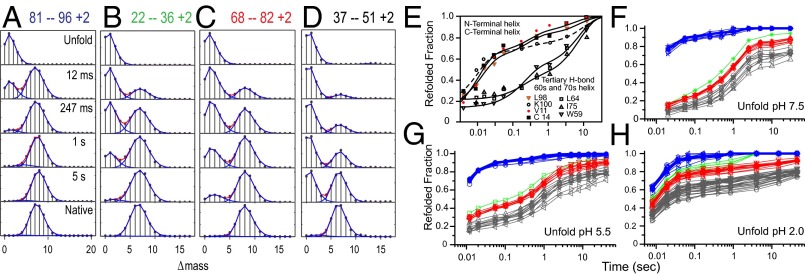
Previous hydrogen exchange (HX) studies of the spontaneous reversible unfolding of Cytochrome c (Cyt c) under native conditions have led to the following conclusions. Native Cyt c (104 residues) is composed of five cooperative folding units, called foldons. The high-energy landscape is dominated by an energy ladder of partially folded forms that differ from each other by one cooperative foldon unit. The reversible equilibrium unfolding of native Cyt c steps up through these intermediate forms to the unfolded state in an energy-ordered sequence, one foldon unit at a time. To more directly study Cyt c intermediates and pathways during normal energetically downhill kinetic folding, the present work used HX pulse labeling analyzed by a fragment separation-mass spectrometry method. The results show that 95% or more of the Cyt c population folds by stepping down through the same set of foldon-dependent pathway intermediates as in energetically uphill equilibrium unfolding. These results add to growing evidence that proteins fold through a classical pathway sequence of native-like intermediates rather than through a vast number of undefinable intermediates and pathways. The present results also emphasize the condition-dependent nature of kinetic barriers, which, with less informative experimental methods (fluorescence, etc.), are often confused with variability in intermediates and pathways.
Keywords: cytochrome c; foldons; protein folding.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Bai Y, Milne JS, Mayne L, Englander SW. Protein stability parameters measured by hydrogen exchange. Proteins. 1994;20(1):4–14. - PubMed
-
- Levinthal C. Are there pathways for protein folding. J Chim Phys. 1968;65:44–45.
-
- Bryngelson JD, Onuchic JN, Socci ND, Wolynes PG. Funnels, pathways, and the energy landscape of protein folding: A synthesis. Proteins. 1995;21(3):167–195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
