

Review
doi: 10.1016/j.cell.2016.02.041.
Hardwiring Stem Cell Communication through Tissue Structure
Affiliations
- PMID: 26967287
- PMCID: PMC4805424
- DOI: 10.1016/j.cell.2016.02.041
Item in Clipboard
Review
Hardwiring Stem Cell Communication through Tissue Structure
Cell.
.
Abstract
Adult stem cells across diverse organs self-renew and differentiate to maintain tissue homeostasis. How stem cells receive input to preserve tissue structure and function largely relies on their communication with surrounding cellular and non-cellular elements. As such, how tissues are organized and patterned not only reflects organ function, but also inherently hardwires networks of communication between stem cells and their environment to direct tissue homeostasis and injury repair. This review highlights how different methods of stem cell communication reflect the unique organization and function of diverse tissues.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
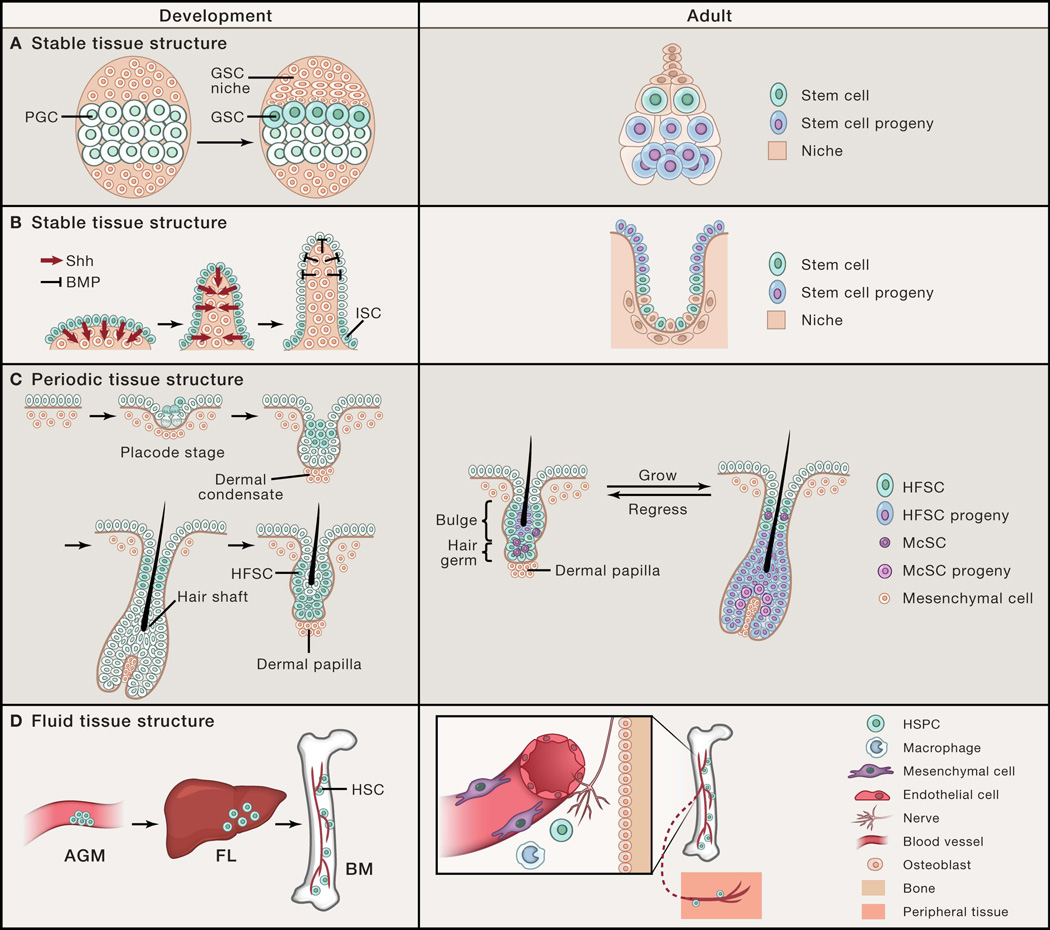
(A) Stable polarized stem cell niche structure in the fly ovary directs asymmetric cell fates between stem cells and their progeny. Drosophila female germline stem cells (GSCs) are specified from primordial germ cells (PGCs) that are adjacent to the newly formed GSC niche during ovary development. In the adult, GSCs are maintained by signals provided by the niche, while displacement of the daughter progeny outside of the niche induces their differentiation. Here, stem cell position is polarized relative to their differentiated progeny and the tissue structure remains stable over time. (B) During intestinal development, intestinal stem cells (ISCs) are restricted to the base of the villi by morphogenesis-induced changes in signaling territories. Similar to the fly ovary, ISC position is polarized relative to their differentiated enterocytes that mobilize upward out of the crypt niche base to villus tip. (C) Hair follicle stem cells (HFSCs) are specified during hair follicle morphogenesis by asymmetric cell divisions and displaced suprabasally. In the adult hair follicle, periodic self-renewal and differentiation of HFSCs and melanocyte stem cells (McSCs) are coordinated with the oscillating hair follicle structure. In this example, the stem cell position is also polarized and fixed; however, the tissue structure periodically changes. (D) Definitive hematopoietic stem cells (HSCs) form in the aorta-gonad-mesonephros (AGM) and migrate to the fetal liver (FL) before populating the bone marrow (BM) during embryogenesis. In the adult, hematopoietic stem and progenitor cells (HSPCs) are centralized in the BM niche, but due to their fluid tissue architecture, can migrate into the blood circulation and travel between peripheral tissues. Here, stem cell position is flexible and mobile and dependent upon dynamic niche interactions.
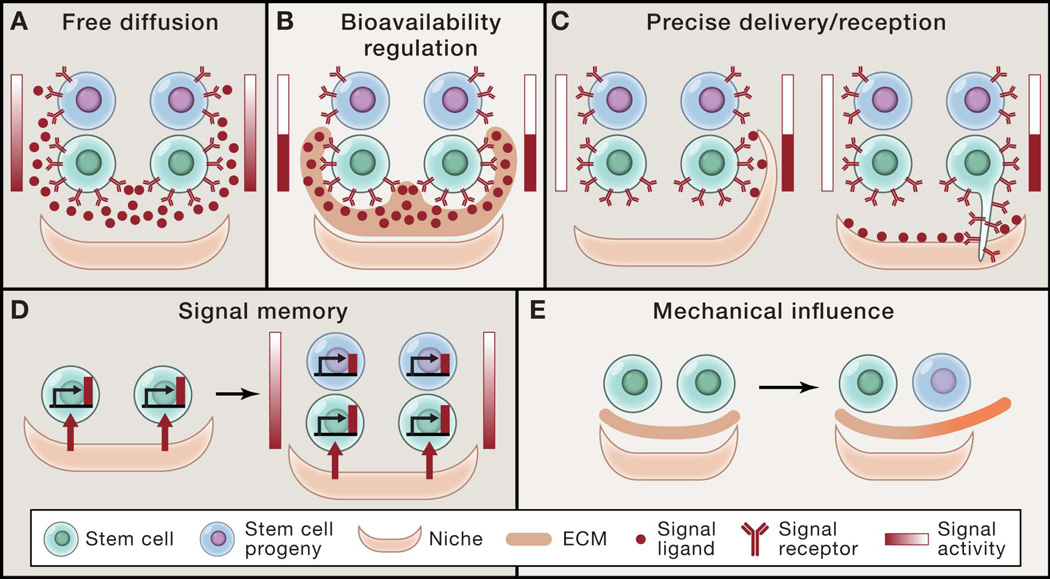
(A) Free diffusion of signal ligand from a polarized source generates a signal gradient across stem cells and their progeny that determines asymmetric cell fates. (B) Regulation of the bioavailability of signal ligand by extracellular matrix (ECM) components can concentrate signal ligand to stem cells, while restricting ligand from differentiated progeny. (C) Specialized sub-cellular structures of either niche cells or stem cells precisely control signal activation in specific stem cells. (D) Differential signal activity between stem cells and their progeny can be generated by signal memory and decay once they lose exposure to the signal source. (E) Changes in mechanical properties such as matrix stiffness can influence stem cell self-renewal and differentiation.
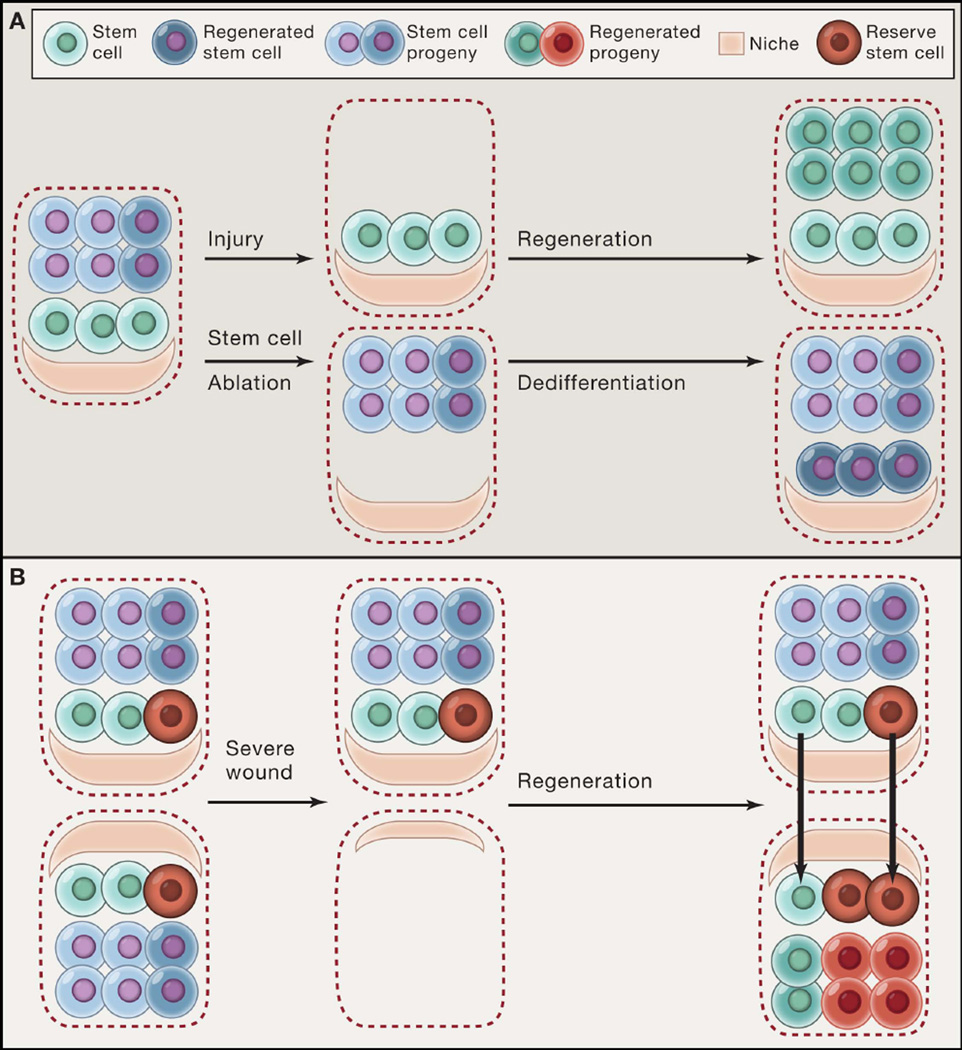
(A) Stem cells regenerate differentiated lineages after injury (e.g. basal stem cells of the upper airway following SO2-mediated damage). However, when stem cells themselves are ablated, their progeny can also dedifferentiate into functional stem cells (e.g. Dll1+ intestinal secretory precursors following irradiation damage). (B) A subset of reserve stem cells (red) that do not normally contribute to homeostasis can be engaged along with other resident stem cells (green) to reconstitute tissue following injury (e.g. K5+p63+ basal stem cells in the distal airway). A recent study suggests that stem cells might regenerate tissue architecture by using the residual niche structures left as a template after severe injury.
References
-
- Adams GB, Chabner KT, Alley IR, Olson DP, Szczepiorkowski ZM, Poznansky MC, Kos CH, Pollak MR, Brown EM, Scadden DT. Stem cell engraftment at the endosteal niche is specified by the calcium-sensing receptor. Nature. 2006;439:599–603. - PubMed
-
- Barker N. Adult intestinal stem cells: critical drivers of epithelial homeostasis and regeneration. Nature reviews Molecular cell biology. 2014;15:19–33. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
