

P66SHC deletion improves fertility and progeric phenotype of late-generation TERC-deficient mice but not their short lifespan
- PMID: 26968134
- PMCID: PMC4854904
- DOI: 10.1111/acel.12448
P66SHC deletion improves fertility and progeric phenotype of late-generation TERC-deficient mice but not their short lifespan
Abstract
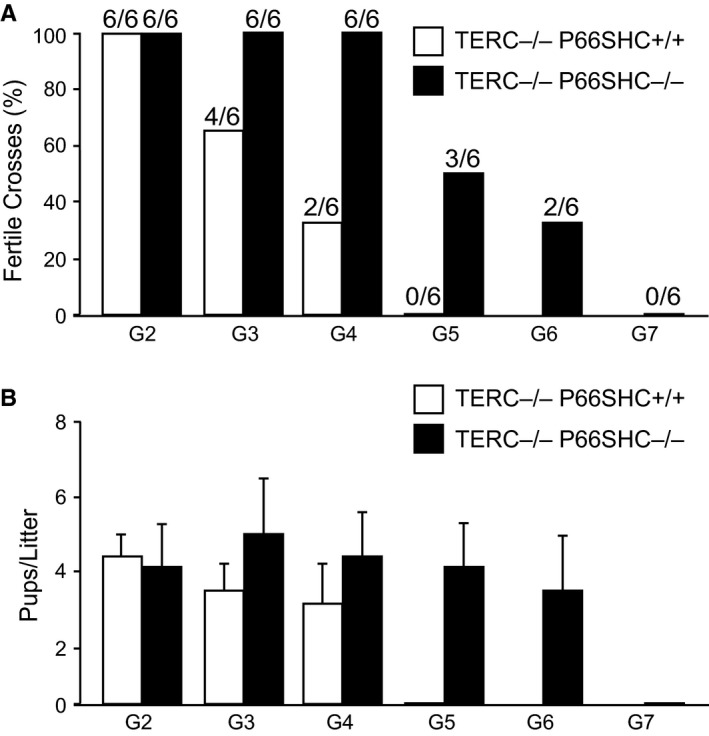
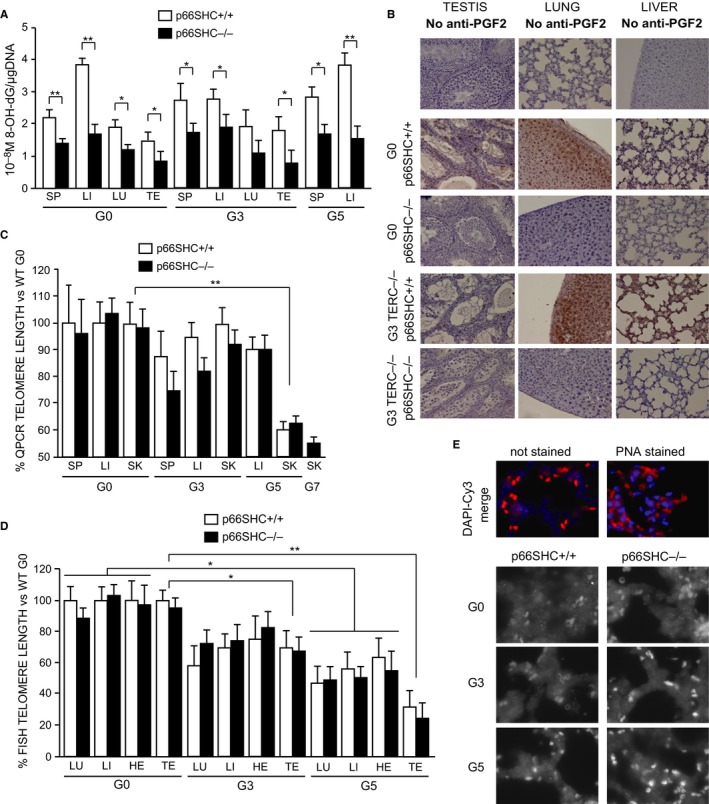
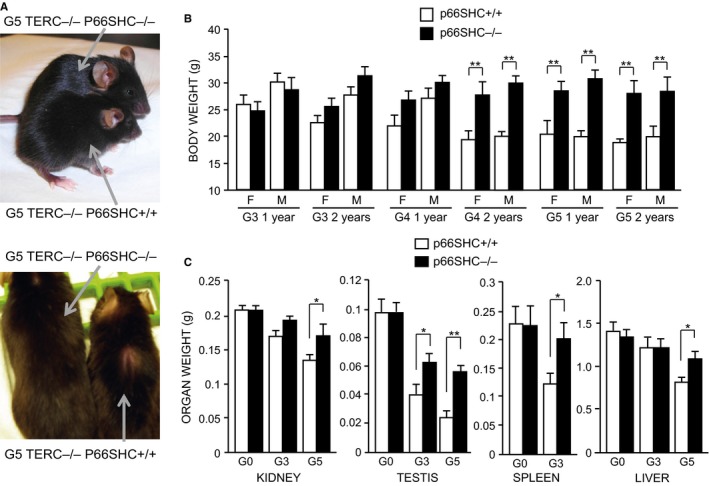
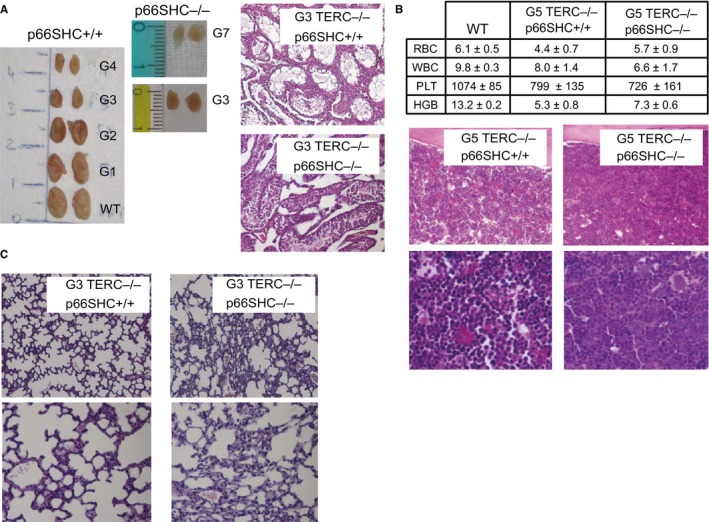
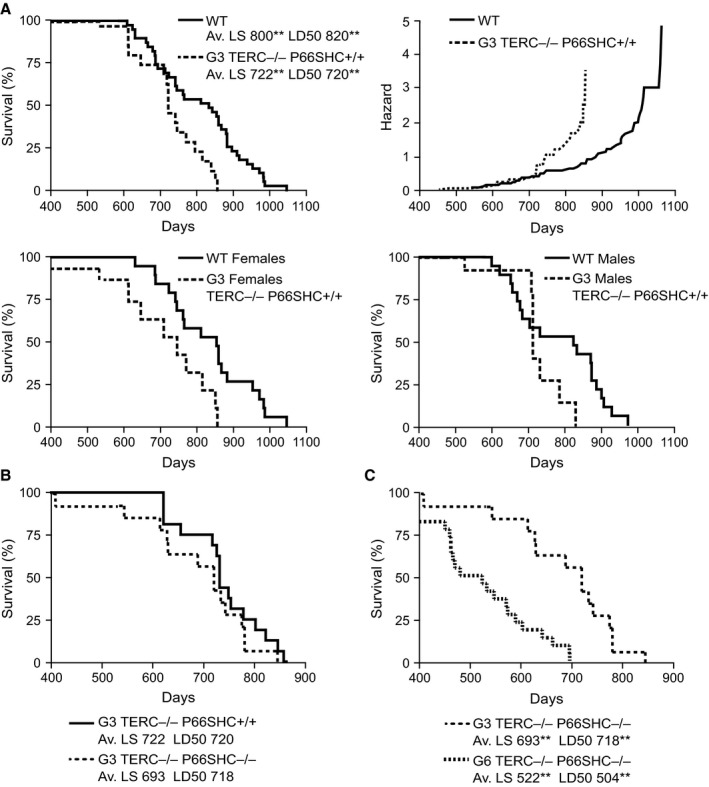
Oxidative stress and telomere attrition are considered the driving factors of aging. As oxidative damage to telomeric DNA favors the erosion of chromosome ends and, in turn, telomere shortening increases the sensitivity to pro-oxidants, these two factors may trigger a detrimental vicious cycle. To check whether limiting oxidative stress slows down telomere shortening and related progeria, we have investigated the effect of p66SHC deletion, which has been shown to reduce oxidative stress and mitochondrial apoptosis, on late-generation TERC (telomerase RNA component)-deficient mice having short telomeres and reduced lifespan. Double mutant (TERC(-/-) p66SHC(-/-) ) mice were generated, and their telomere length, fertility, and lifespan investigated in different generations. Results revealed that p66SHC deletion partially rescues sterility and weight loss, as well as organ atrophy, of TERC-deficient mice, but not their short lifespan and telomere erosion. Therefore, our data suggest that p66SHC-mediated oxidative stress and telomere shortening synergize in some tissues (including testes) to accelerate aging; however, early mortality of late-generation mice seems to be independent of any link between p66SHC-mediated oxidative stress and telomere attrition.
Keywords: fertility; lifespan; oxidative DNA damage; telo-meres.
© 2016 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Figures

References
-
- Albiero M, Poncina N, Tjwa M, Ciciliot S, Menegazzo L, Ceolotto G, Vigili de Kreutzenberg S, Moura R, Giorgio M, Pelicci P, Avogaro A, Fadini GP (2014) Diabetes causes bone marrow autonomic neuropathy and impairs stem cell mobilization via dysregulated p66Shc and Sirt1. Diabetes 63, 1353–1365. - PubMed
-
- Berry A, Capone F, Giorgio M, Pelicci PG, de Kloet ER, Alleva E, Minghetti L, Cirulli F (2007) Deletion of the life span determinant p66Shc prevents age‐dependent increases in emotionality and pain sensitivity in mice. Exp. Gerontol. 422, 37–45. - PubMed
-
- Blackburn EH, Greider CW, Szostak JW (2006) Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. Nat. Med. 12, 1133–1138. - PubMed
-
- Carpi A, Menabò R, Kaludercic N, Pelicci P, Di Lisa F, Giorgio M (2009) The cardioprotective effects elicited by p66(Shc) ablation demonstrate the crucial role of mitochondrial ROS formation in ischemia/reperfusion injury. Biochim. Biophys. Acta 1787, 774–780. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
