

Temperature response of bundle-sheath conductance in maize leaves
- PMID: 26969744
- PMCID: PMC4861018
- DOI: 10.1093/jxb/erw104
Temperature response of bundle-sheath conductance in maize leaves
Abstract
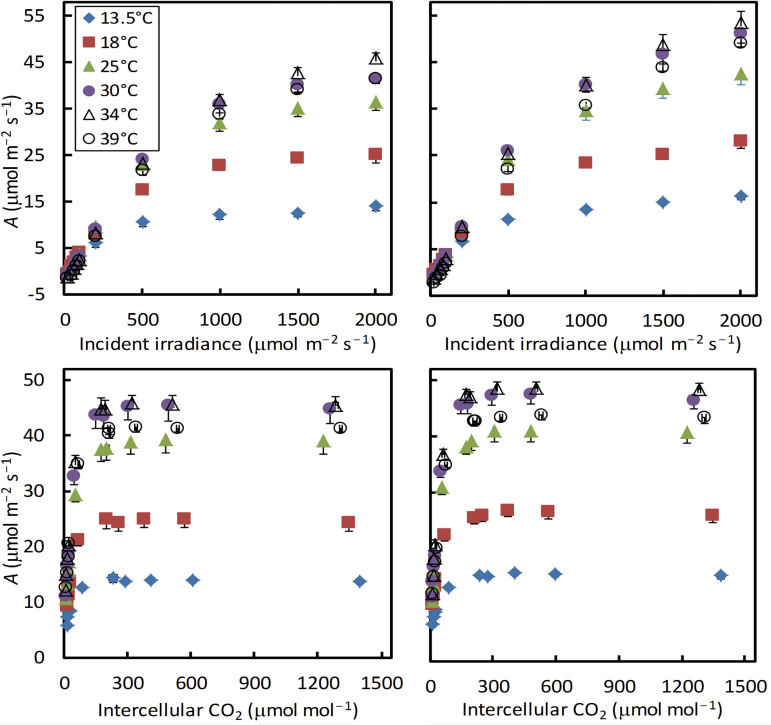
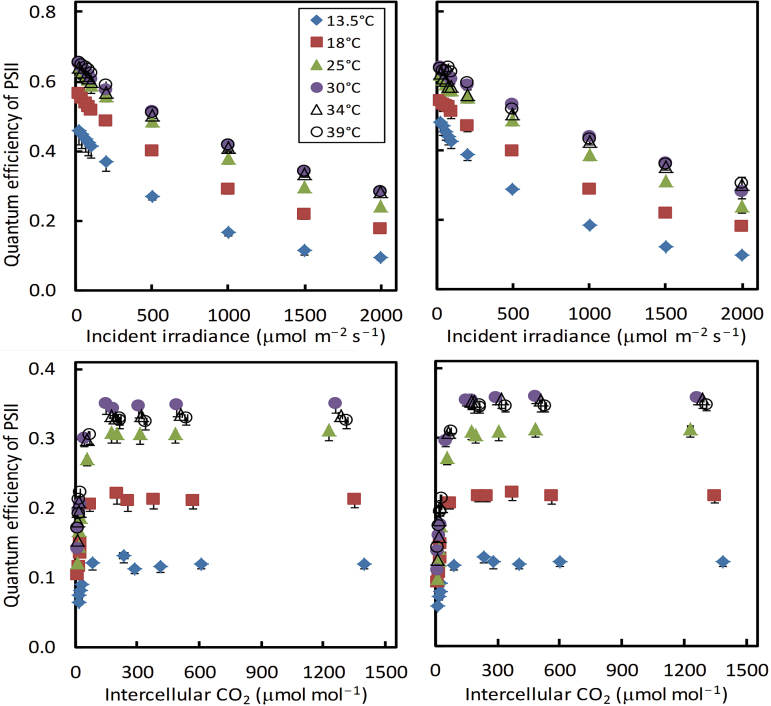
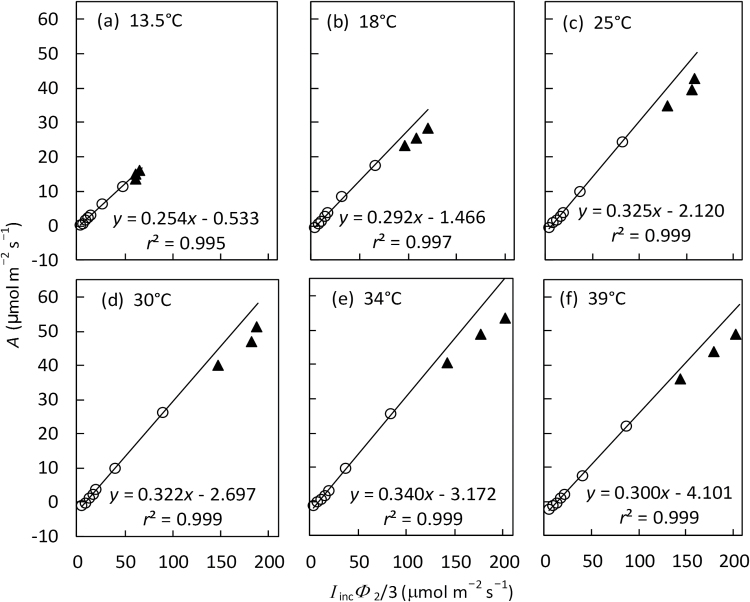
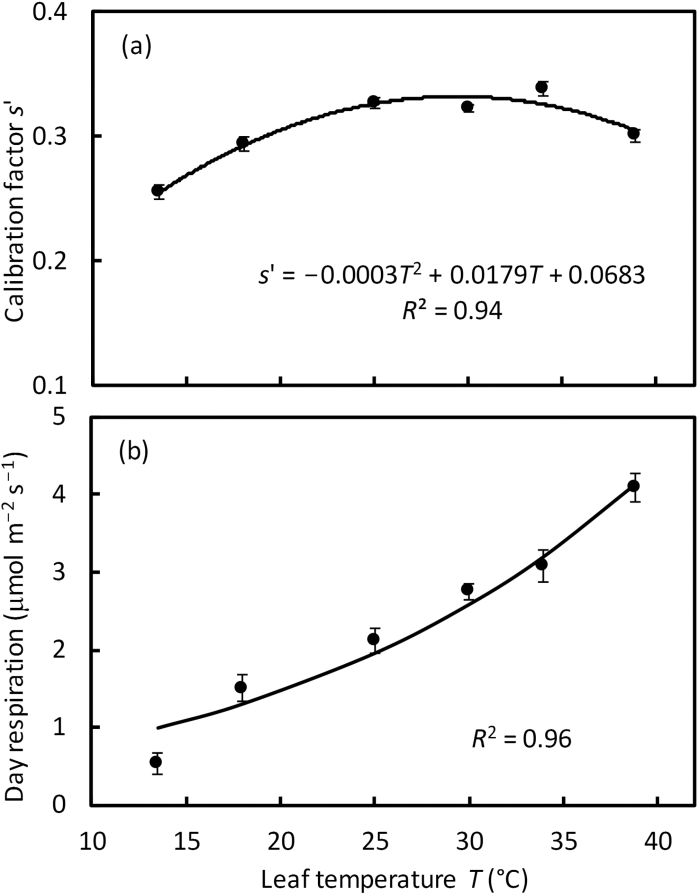
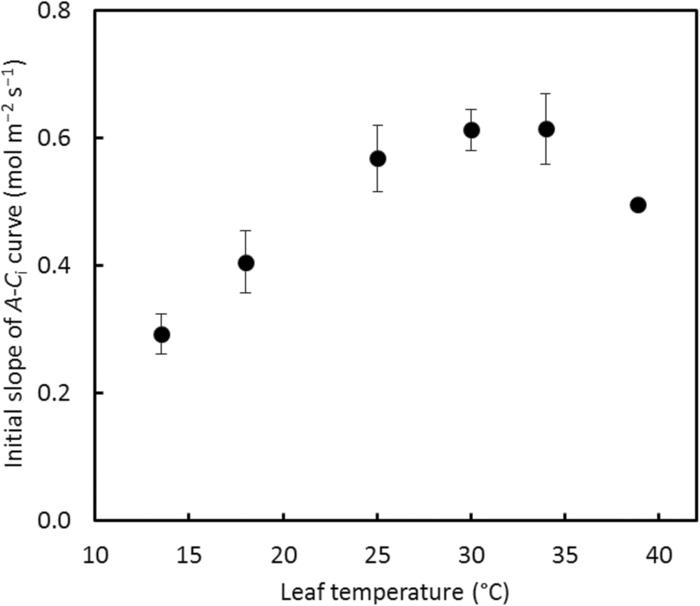
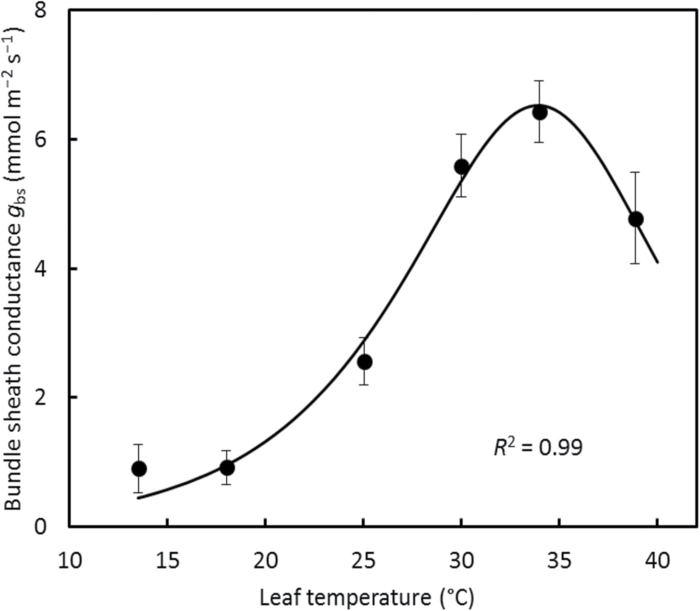
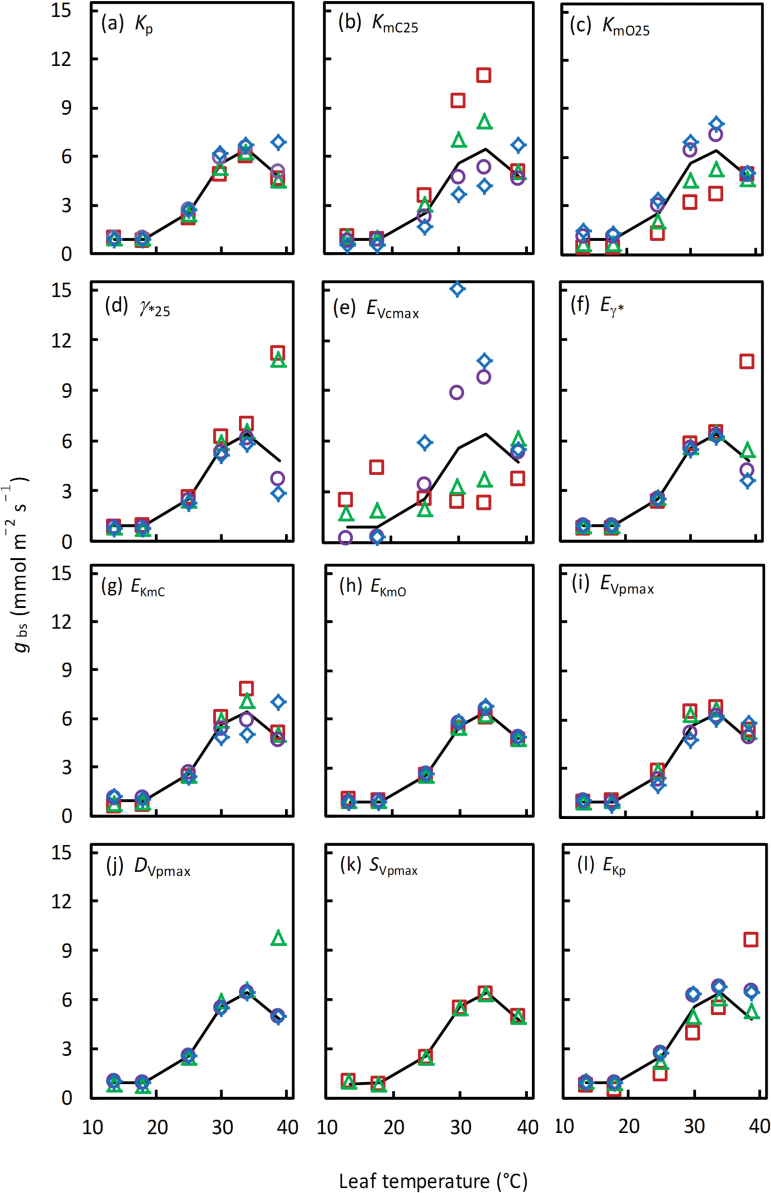
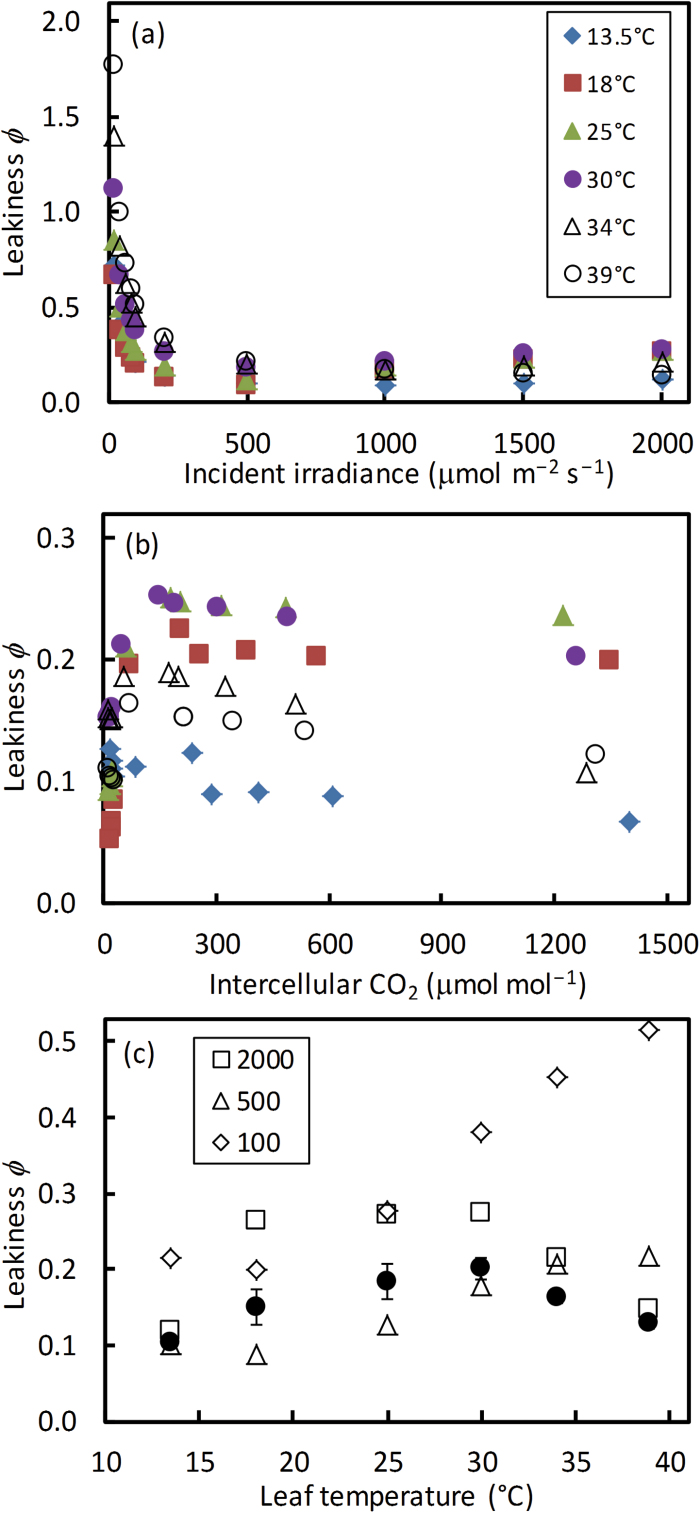
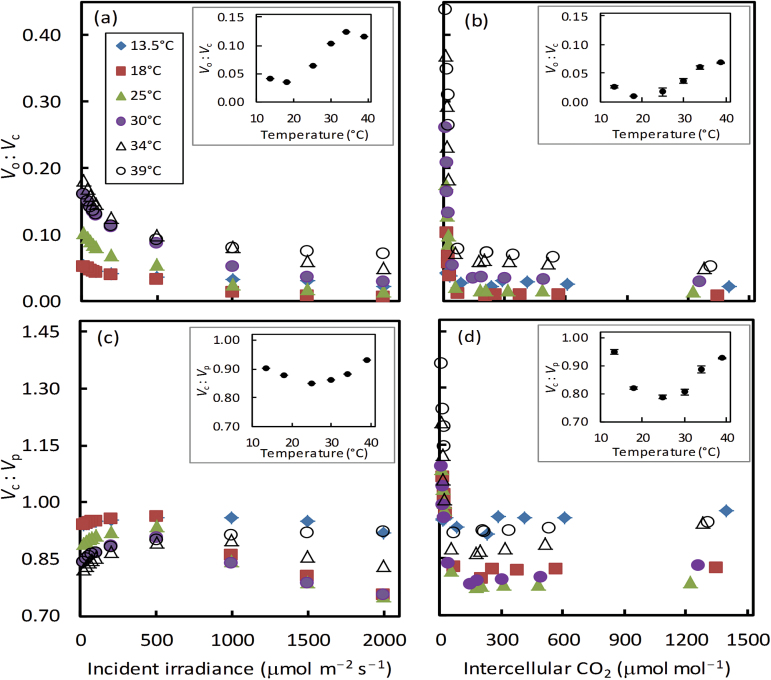
A small bundle-sheath conductance (g bs) is essential for the C4 CO2-concentrating mechanism to suppress photorespiration effectively. To predict the productivity of C4 crops accurately under global warming, it is necessary to examine whether and how g bs responds to temperature. We investigated the temperature response of g bs in maize by fitting a C4 photosynthesis model to combined gas exchange and chlorophyll fluorescence measurements of irradiance and CO2 response curves at 21% and 2% O2 within the range of 13.5-39 °C. The analysis was based on reported kinetic constants of C4 Rubisco and phosphoenolpyruvate carboxylase and temperature responses of C3 mesophyll conductance (g m). The estimates of g bs varied greatly with leaf temperature. The temperature response of g bs was well described by the peaked Arrhenius equation, with the optimum temperature being ~34 °C. The assumed temperature responses of g m had only a slight impact on the temperature response of g bs In contrast, using extreme values of some enzyme kinetic constants changed the shape of the response, from the peaked optimum response to the non-peaked Arrhenius pattern. Further studies are needed to confirm such an Arrhenius response pattern from independent measurement techniques and to assess whether it is common across C4 species.
Keywords: Diffusive resistance; Zea mays.; maximum PEPc activity; maximum Rubisco activity; modelling; warming effect.
© The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Badger MR, Collatz GJ. 1977. Studies on the kinetic mechanism of ribulose-1,5-biophosphate carboxylase and oxygenase reactions, with particular reference to the effect of temperature on kinetic parameters. Carnegie Instute of Washington Yearbook 76, 355–361.
-
- Badger MR, Price GD. 1994. The role of carbonic anhydrase in photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 45, 369–392.
-
- Barbour MM, Evans JR, Simonin KA, von Caemmerer S. 2016. Online CO2 and H2O oxygen isotope fractionation allows estimation of mesophyll conductance in C4 plants, and reveals that mesophyll conductance decreases as leaves age in both C4 and C3 plants. New Phytologist (in press). - PubMed
-
- Bellasio C, Griffiths H. 2013. Acclimation to low light by C4 maize: implications for bundle sheath leakiness. Plant, Cell and Environment 37, 1046–1058. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
