

Mapping and modeling of a strain-specific epitope in the Norwalk virus capsid inner shell
- PMID: 26971245
- PMCID: PMC11036327
- DOI: 10.1016/j.virol.2016.02.019
Mapping and modeling of a strain-specific epitope in the Norwalk virus capsid inner shell
Abstract
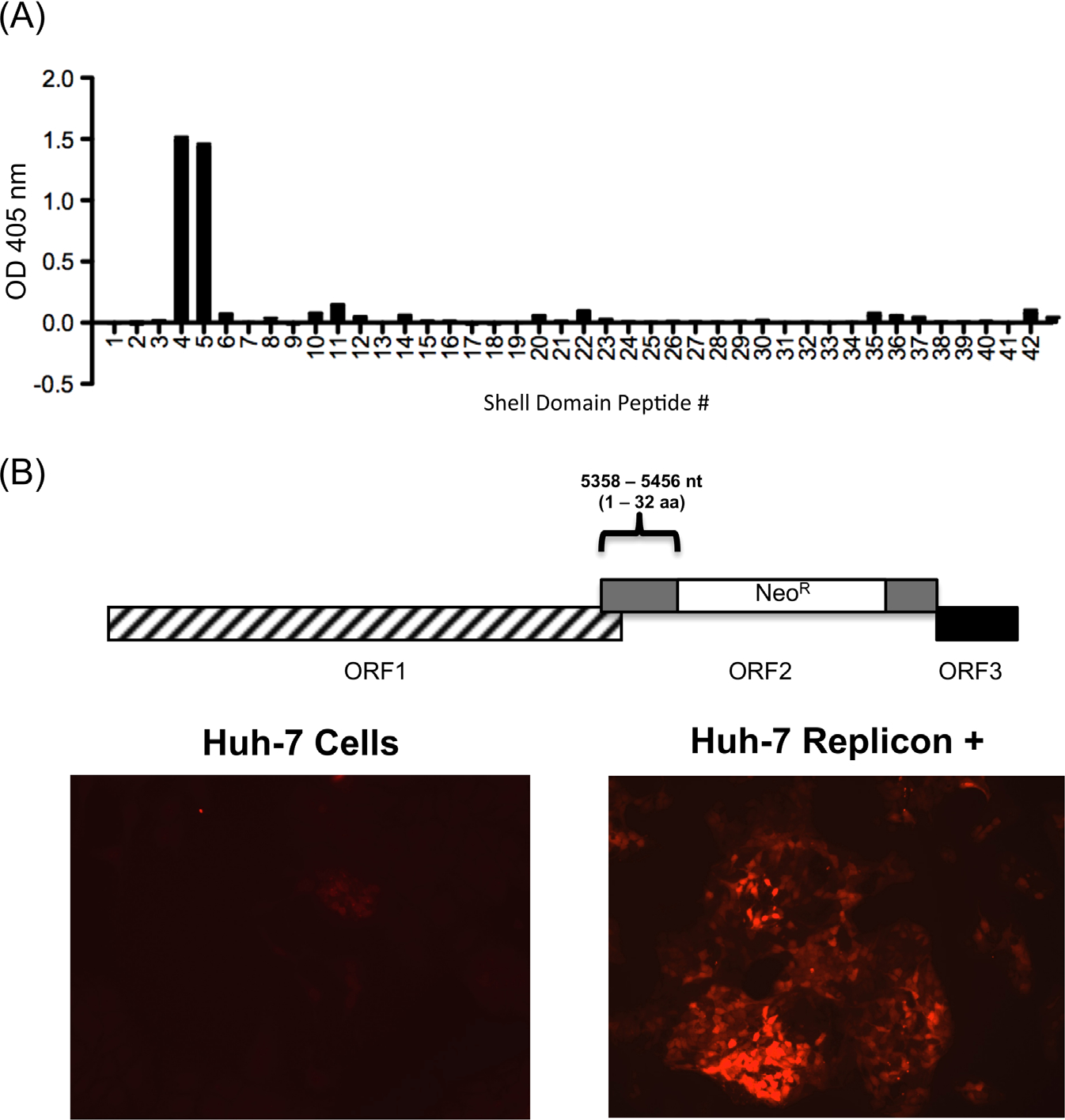
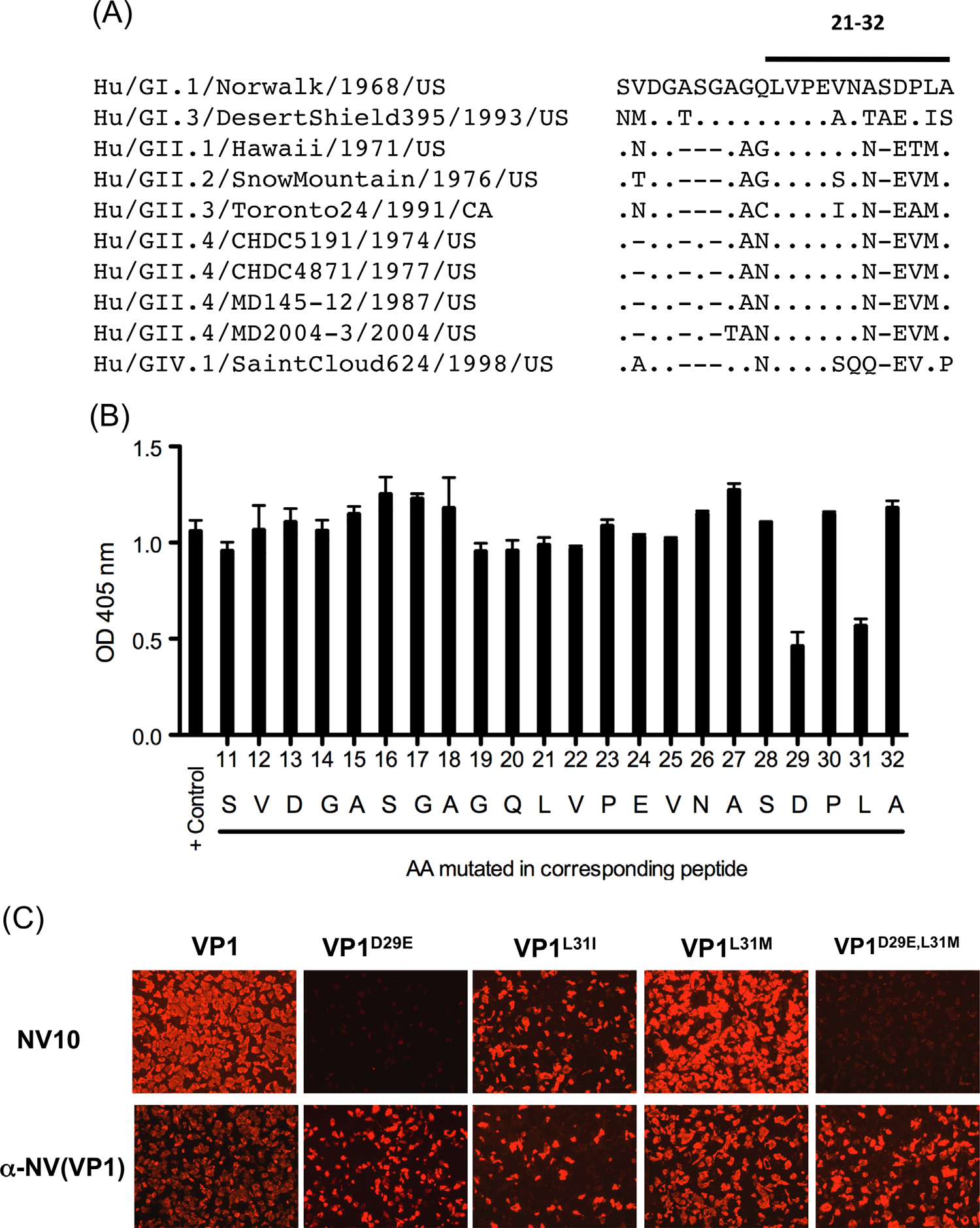
Noroviruses are diverse positive-strand RNA viruses associated with acute gastroenteritis. Cross-reactive epitopes have been mapped primarily to conserved sequences in the capsid VP1 Shell (S) domain, and strain-specific epitopes to the highly variable Protruding (P) domain. In this work, we investigated a strain-specific linear epitope defined by MAb NV10 that was raised against prototype (Genogroup I.1) strain Norwalk virus (NV). Using peptide scanning and mutagenesis, the epitope was mapped to amino acids 21-32 (LVPEVNASDPLA) of the NV S domain, and its specificity was verified by epitope transfer and reactivity with a recombinant MAb NV10 single-chain variable fragment (scFv). Comparative structural modeling of the NV10 strain-specific and the broadly cross-reactive TV20 epitopes identified two internal non-overlapping sites in the NV shell, corresponding to variable and conserved amino acid sequences among strains, respectively. The S domain, like the P domain, contains strain-specific epitopes that contribute to the antigenic diversity among the noroviruses.
Keywords: Caliciviruses; Epitope-tagging; Noroviruses.
Published by Elsevier Inc.
Figures




Similar articles
-
The Antigenic Topology of Norovirus as Defined by B and T Cell Epitope Mapping: Implications for Universal Vaccines and Therapeutics.Viruses. 2019 May 10;11(5):432. doi: 10.3390/v11050432. Viruses. 2019. PMID: 31083353 Free PMC article. Review.
-
Characterization of monoclonal antibodies generated against Norwalk virus GII capsid protein expressed in Escherichia coli.Microbiol Immunol. 2000;44(11):905-14. doi: 10.1111/j.1348-0421.2000.tb02582.x. Microbiol Immunol. 2000. PMID: 11145271
-
Identification of a Broadly Cross-Reactive Epitope in the Inner Shell of the Norovirus Capsid.PLoS One. 2013 Jun 21;8(6):e67592. doi: 10.1371/journal.pone.0067592. Print 2013. PLoS One. 2013. PMID: 23805319 Free PMC article.
-
Characterization of a cross-reactive linear epitope in human genogroup I and bovine genogroup III norovirus capsid proteins.Virology. 2006 Dec 5-20;356(1-2):179-87. doi: 10.1016/j.virol.2006.07.034. Epub 2006 Aug 24. Virology. 2006. PMID: 16934306
-
Antigenic mapping of the recombinant Norwalk virus capsid protein using monoclonal antibodies.Virology. 1996 Mar 1;217(1):252-61. doi: 10.1006/viro.1996.0112. Virology. 1996. PMID: 8599210
Cited by
-
Identification of a novel fully human anti-toxic shock syndrome toxin (TSST)-1 single-chain variable fragment antibody averting TSST-1-induced mitogenesis and cytokine secretion.BMC Biotechnol. 2022 Oct 28;22(1):31. doi: 10.1186/s12896-022-00760-8. BMC Biotechnol. 2022. PMID: 36307814 Free PMC article.
-
The Antigenic Topology of Norovirus as Defined by B and T Cell Epitope Mapping: Implications for Universal Vaccines and Therapeutics.Viruses. 2019 May 10;11(5):432. doi: 10.3390/v11050432. Viruses. 2019. PMID: 31083353 Free PMC article. Review.
-
High-Resolution Mapping of Human Norovirus Antigens via Genomic Phage Display Library Selections and Deep Sequencing.J Virol. 2020 Dec 9;95(1):e01495-20. doi: 10.1128/JVI.01495-20. Print 2020 Dec 9. J Virol. 2020. PMID: 33055250 Free PMC article.
-
Bat Caliciviruses and Human Noroviruses Are Antigenically Similar and Have Overlapping Histo-Blood Group Antigen Binding Profiles.mBio. 2018 May 22;9(3):e00869-18. doi: 10.1128/mBio.00869-18. mBio. 2018. PMID: 29789360 Free PMC article.
-
Broad-range and effective detection of human noroviruses by colloidal gold immunochromatographic assay based on the shell domain of the major capsid protein.BMC Microbiol. 2021 Jan 11;21(1):22. doi: 10.1186/s12866-020-02084-z. BMC Microbiol. 2021. PMID: 33430771 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
