

Candida albicans repetitive elements display epigenetic diversity and plasticity
- PMID: 26971880
- PMCID: PMC4789652
- DOI: 10.1038/srep22989
Candida albicans repetitive elements display epigenetic diversity and plasticity
Abstract
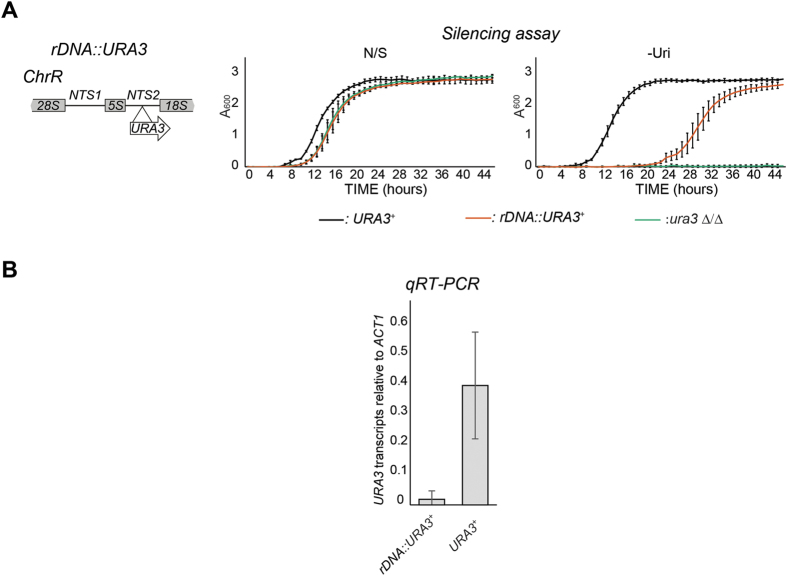
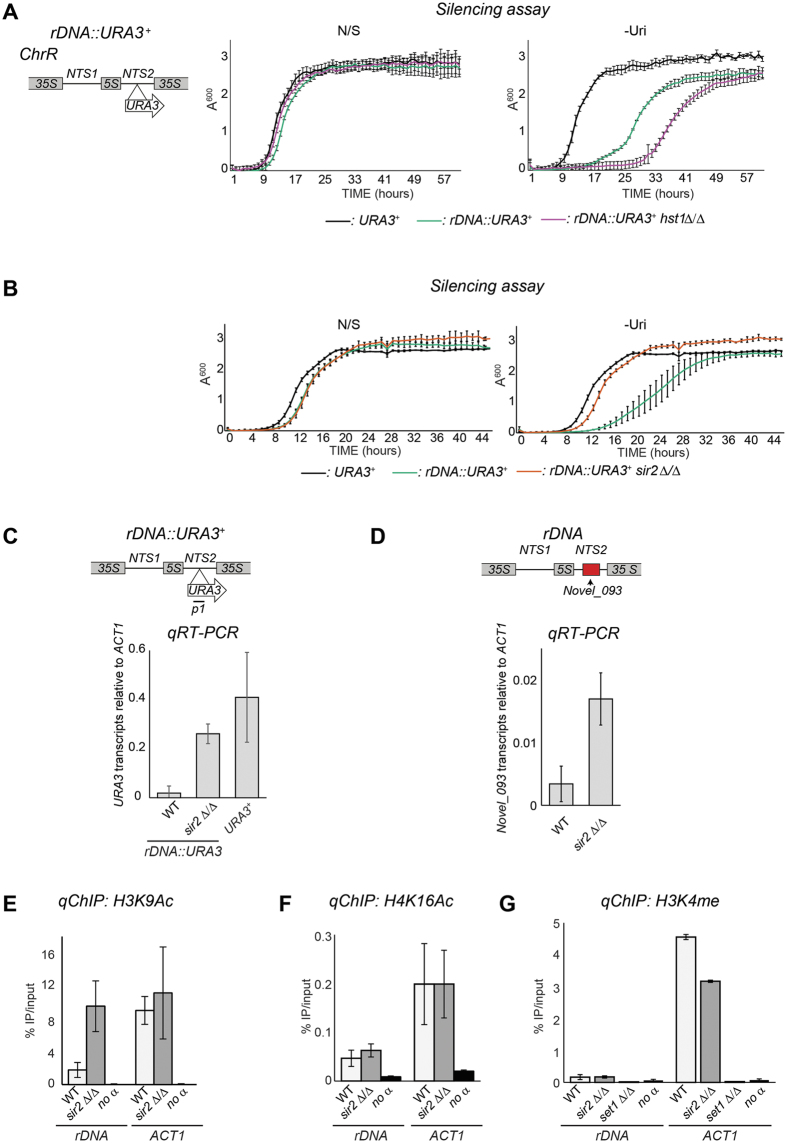
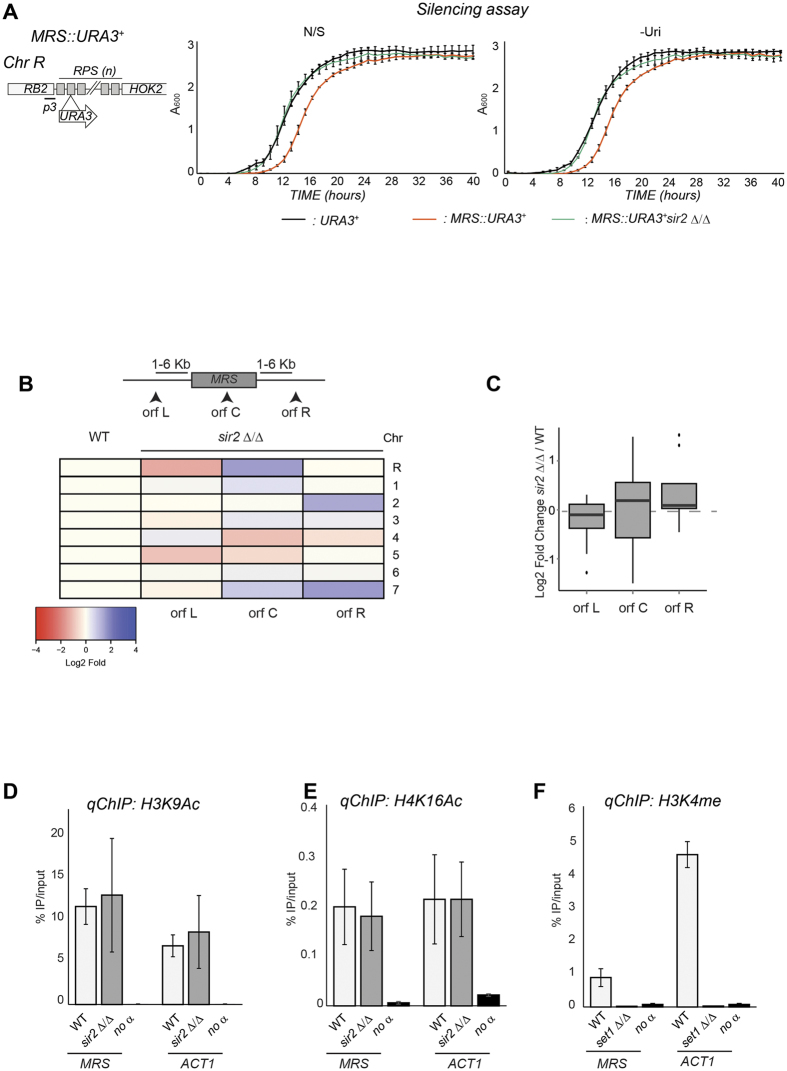
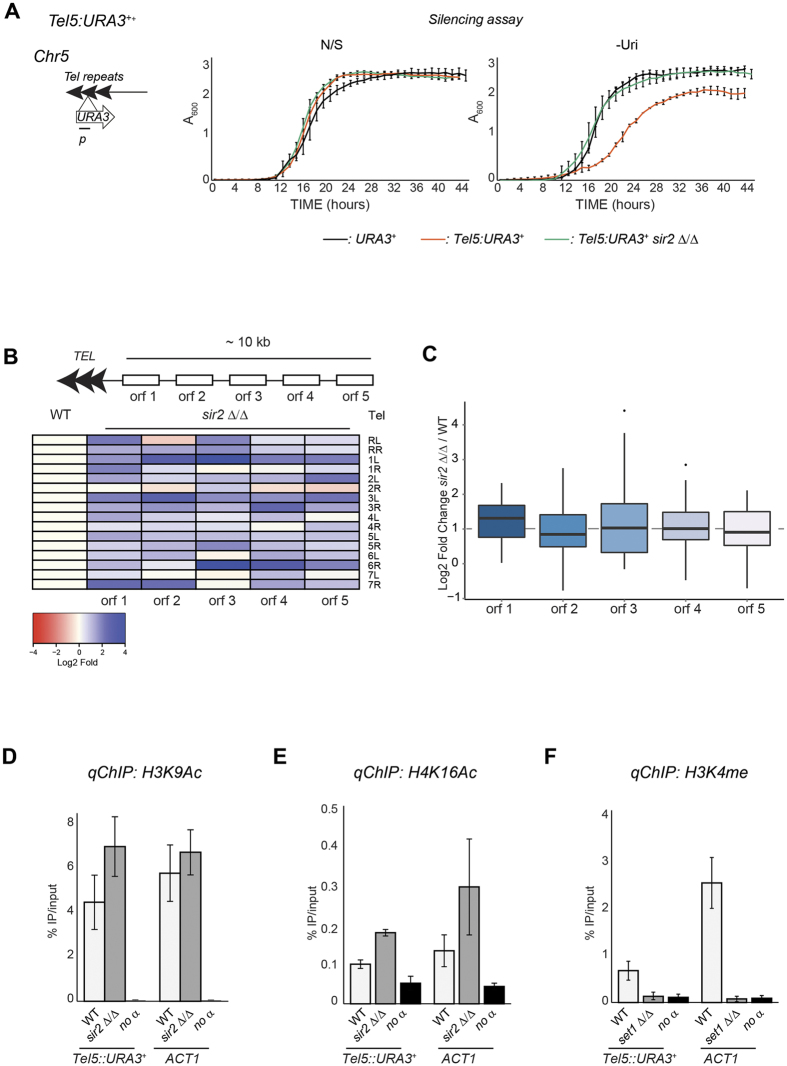
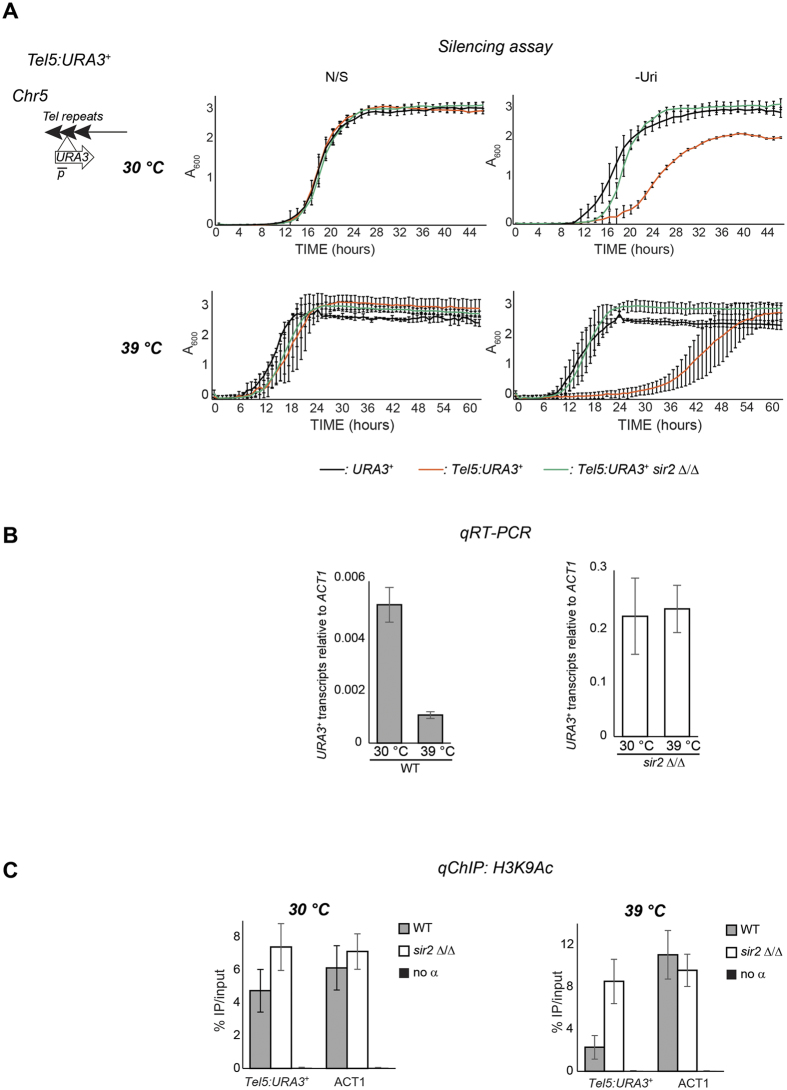
Transcriptionally silent heterochromatin is associated with repetitive DNA. It is poorly understood whether and how heterochromatin differs between different organisms and whether its structure can be remodelled in response to environmental signals. Here, we address this question by analysing the chromatin state associated with DNA repeats in the human fungal pathogen Candida albicans. Our analyses indicate that, contrary to model systems, each type of repetitive element is assembled into a distinct chromatin state. Classical Sir2-dependent hypoacetylated and hypomethylated chromatin is associated with the rDNA locus while telomeric regions are assembled into a weak heterochromatin that is only mildly hypoacetylated and hypomethylated. Major Repeat Sequences, a class of tandem repeats, are assembled into an intermediate chromatin state bearing features of both euchromatin and heterochromatin. Marker gene silencing assays and genome-wide RNA sequencing reveals that C. albicans heterochromatin represses expression of repeat-associated coding and non-coding RNAs. We find that telomeric heterochromatin is dynamic and remodelled upon an environmental change. Weak heterochromatin is associated with telomeres at 30 °C, while robust heterochromatin is assembled over these regions at 39 °C, a temperature mimicking moderate fever in the host. Thus in C. albicans, differential chromatin states controls gene expression and epigenetic plasticity is linked to adaptation.
Figures
Similar articles
-
Chromatin Profiling of the Repetitive and Nonrepetitive Genomes of the Human Fungal Pathogen Candida albicans.mBio. 2019 Jul 23;10(4):e01376-19. doi: 10.1128/mBio.01376-19. mBio. 2019. PMID: 31337722 Free PMC article.
-
To Repeat or Not to Repeat: Repetitive Sequences Regulate Genome Stability in Candida albicans.Genes (Basel). 2019 Oct 30;10(11):866. doi: 10.3390/genes10110866. Genes (Basel). 2019. PMID: 31671659 Free PMC article. Review.
-
Differential association of Orc1 and Sir2 proteins to telomeric domains in Plasmodium falciparum.J Cell Sci. 2008 Jun 15;121(Pt 12):2046-53. doi: 10.1242/jcs.026427. J Cell Sci. 2008. PMID: 18525026
-
Epigenetic analyses and the distribution of repetitive DNA and resistance genes reveal the complexity of common bean (Phaseolus vulgaris L., Fabaceae) heterochromatin.Cytogenet Genome Res. 2014;143(1-3):168-78. doi: 10.1159/000360572. Epub 2014 Apr 15. Cytogenet Genome Res. 2014. PMID: 24752176
-
RNA meets chromatin.Genes Dev. 2005 Jul 15;19(14):1635-55. doi: 10.1101/gad.1324305. Genes Dev. 2005. PMID: 16024654 Review.
Cited by
-
The Chromatin of Candida albicans Pericentromeres Bears Features of Both Euchromatin and Heterochromatin.Front Microbiol. 2016 May 19;7:759. doi: 10.3389/fmicb.2016.00759. eCollection 2016. Front Microbiol. 2016. PMID: 27242771 Free PMC article.
-
Sir2 regulates stability of repetitive domains differentially in the human fungal pathogen Candida albicans.Nucleic Acids Res. 2016 Nov 2;44(19):9166-9179. doi: 10.1093/nar/gkw594. Epub 2016 Jul 1. Nucleic Acids Res. 2016. PMID: 27369382 Free PMC article.
-
Genome plasticity in Candida albicans is driven by long repeat sequences.Elife. 2019 Jun 7;8:e45954. doi: 10.7554/eLife.45954. Elife. 2019. PMID: 31172944 Free PMC article.
-
Within-Host Genomic Diversity of Candida albicans in Healthy Carriers.Sci Rep. 2019 Feb 22;9(1):2563. doi: 10.1038/s41598-019-38768-4. Sci Rep. 2019. PMID: 30796326 Free PMC article.
-
Sporadic Gene Loss After Duplication Is Associated with Functional Divergence of Sirtuin Deacetylases Among Candida Yeast Species.G3 (Bethesda). 2016 Oct 13;6(10):3297-3305. doi: 10.1534/g3.116.033845. G3 (Bethesda). 2016. PMID: 27543294 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
