

Distinct Contribution of Adult-Born Hippocampal Granule Cells to Context Encoding
- PMID: 26971949
- PMCID: PMC4962695
- DOI: 10.1016/j.neuron.2016.02.019
Distinct Contribution of Adult-Born Hippocampal Granule Cells to Context Encoding
Abstract
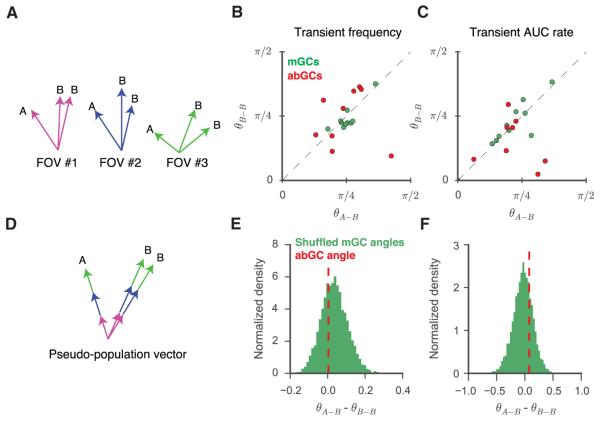
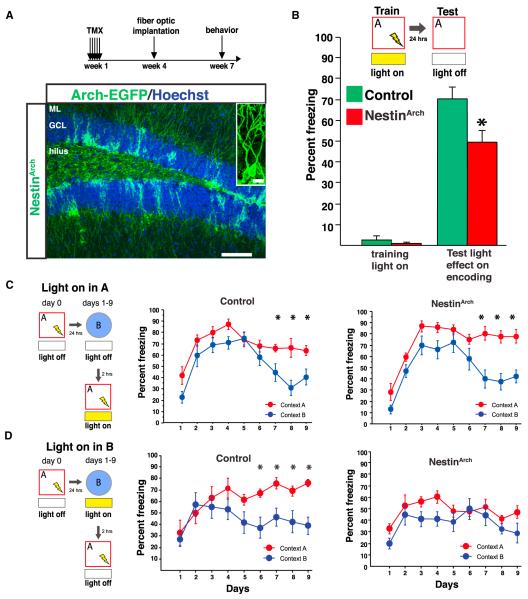
Adult-born granule cells (abGCs) have been implicated in cognition and mood; however, it remains unknown how these cells behave in vivo. Here, we have used two-photon calcium imaging to monitor the activity of young abGCs in awake behaving mice. We find that young adult-born neurons fire at a higher rate in vivo but paradoxically exhibit less spatial tuning than their mature counterparts. When presented with different contexts, mature granule cells underwent robust remapping of their spatial representations, and the few spatially tuned adult-born cells remapped to a similar degree. We next used optogenetic silencing to confirm the direct involvement of abGCs in context encoding and discrimination, consistent with their proposed role in pattern separation. These results provide the first in vivo characterization of abGCs and reveal their participation in the encoding of novel information.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
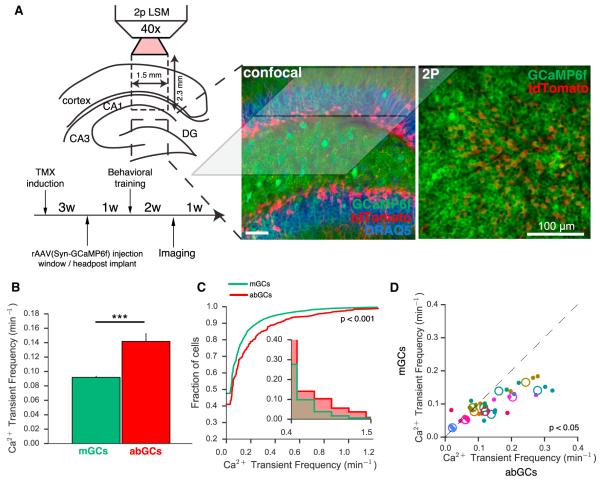
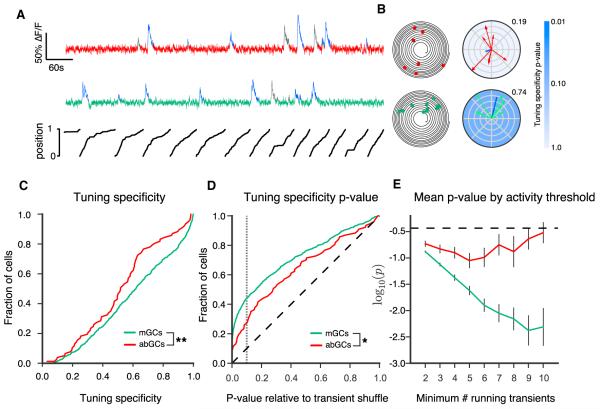
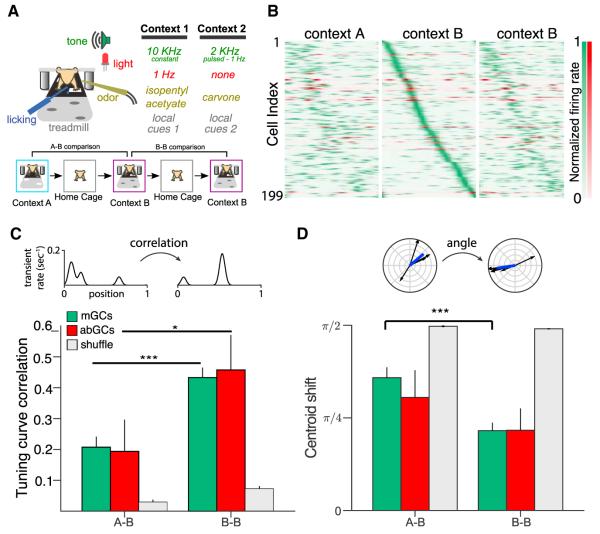
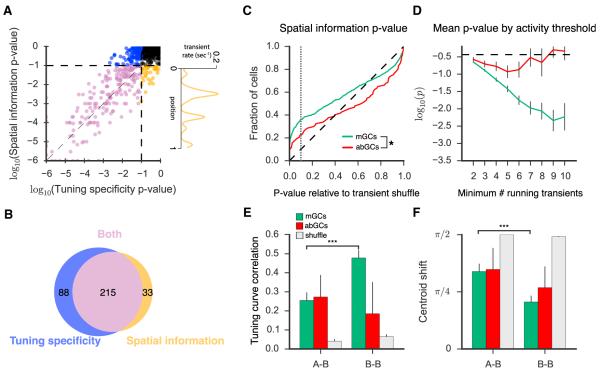
Comment in
-
The Young and the Promiscuous.Neuron. 2016 Apr 6;90(1):6-8. doi: 10.1016/j.neuron.2016.03.021. Neuron. 2016. PMID: 27054613
References
-
- Alme CB, Buzzetti RA, Marrone DF, Leutgeb JK, Chawla MK, Schaner MJ, Bohanick JD, Khoboko T, Leutgeb S, Moser EI, et al. Hippocampal granule cells opt for early retirement. Hippocampus. 2010;20:1109–1123. - PubMed
-
- Bonnevie T, Dunn B, Fyhn M, Hafting T, Derdikman D, Kubie JL, Roudi Y, Moser EI, Moser MB. Grid cells require excitatory drive from the hippocampus. Nat. Neurosci. 2013;16:309–317. - PubMed
-
- Brandt MD, Jessberger S, Steiner B, Kronenberg G, Reuter K, Bick-Sander A, von der Behrens W, Kempermann G. Transient calretinin expression defines early postmitotic step of neuronal differentiation in adult hippocampal neurogenesis of mice. Mol. Cell. Neurosci. 2003;24:603–613. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 1U01NS090583/NS/NINDS NIH HHS/United States
- 1R01MH100631/MH/NIMH NIH HHS/United States
- F30NS090819/NS/NINDS NIH HHS/United States
- R01 MH083862/MH/NIMH NIH HHS/United States
- K01MH099371/MH/NIMH NIH HHS/United States
- R01 MH100631/MH/NIMH NIH HHS/United States
- R01 NS094668/NS/NINDS NIH HHS/United States
- R01 AG043688/AG/NIA NIH HHS/United States
- 1R01NS094668/NS/NINDS NIH HHS/United States
- R37 MH068542/MH/NIMH NIH HHS/United States
- F30 NS090819/NS/NINDS NIH HHS/United States
- K01 MH099371/MH/NIMH NIH HHS/United States
- T32 MH015174/MH/NIMH NIH HHS/United States
- U01 NS090583/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
