

A porcine model of osteosarcoma
- PMID: 26974205
- PMCID: PMC4815050
- DOI: 10.1038/oncsis.2016.19
A porcine model of osteosarcoma
Abstract
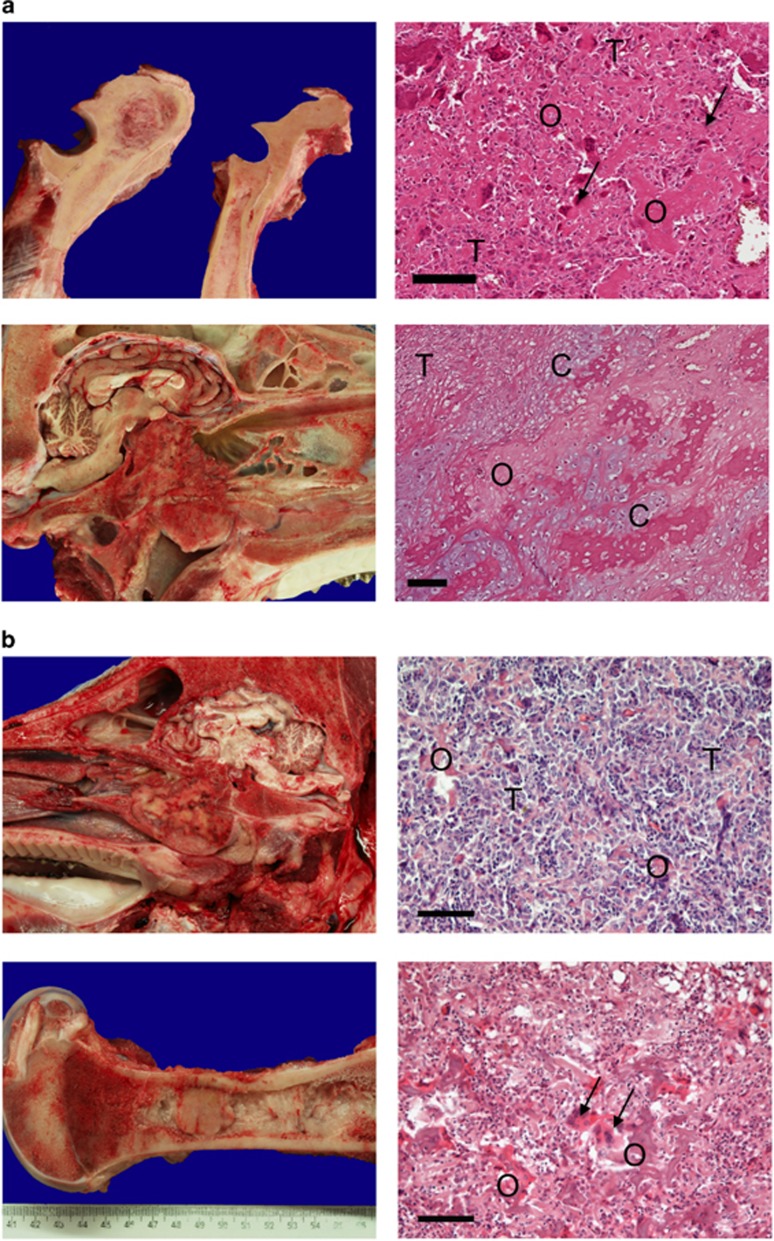
We previously produced pigs with a latent oncogenic TP53 mutation. Humans with TP53 germline mutations are predisposed to a wide spectrum of early-onset cancers, predominantly breast, brain, adrenal gland cancer, soft tissue sarcomas and osteosarcomas. Loss of p53 function has been observed in >50% of human cancers. Here we demonstrate that porcine mesenchymal stem cells (MSCs) convert to a transformed phenotype after activation of latent oncogenic TP53(R167H) and KRAS(G12D), and overexpression of MYC promotes tumorigenesis. The process mimics key molecular aspects of human sarcomagenesis. Transformed porcine MSCs exhibit genomic instability, with complex karyotypes, and develop into sarcomas on transplantation into immune-deficient mice. In pigs, heterozygous knockout of TP53 was sufficient for spontaneous osteosarcoma development in older animals, whereas homozygous TP53 knockout resulted in multiple large osteosarcomas in 7-8-month-old animals. This is the first report that engineered mutation of an endogenous tumour-suppressor gene leads to invasive cancer in pigs. Unlike in Trp53 mutant mice, osteosarcoma developed in the long bones and skull, closely recapitulating the human disease. These animals thus promise a model for juvenile osteosarcoma, a relatively uncommon but devastating disease.
Figures





References
-
- Rangarajan A, Hong SJ, Gifford A, Weinberg RA. Species- and cell type-specific requirements for cellular transformation. Cancer Cell 2004; 6: 171–183. - PubMed
-
- Kendall SD, Linardic CM, Adam SJ, Counter CM. A network of genetic events sufficient to convert normal human cells to a tumorigenic state. Cancer Res 2005; 65: 9824–9828. - PubMed
-
- Flisikowska T, Kind A, Schnieke A. Genetically modified pigs to model human diseases. J Appl Genet 2013; 55: 53–64. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
