

Appetite controlled by a cholecystokinin nucleus of the solitary tract to hypothalamus neurocircuit
- PMID: 26974347
- PMCID: PMC4861598
- DOI: 10.7554/eLife.12225
Appetite controlled by a cholecystokinin nucleus of the solitary tract to hypothalamus neurocircuit
Abstract
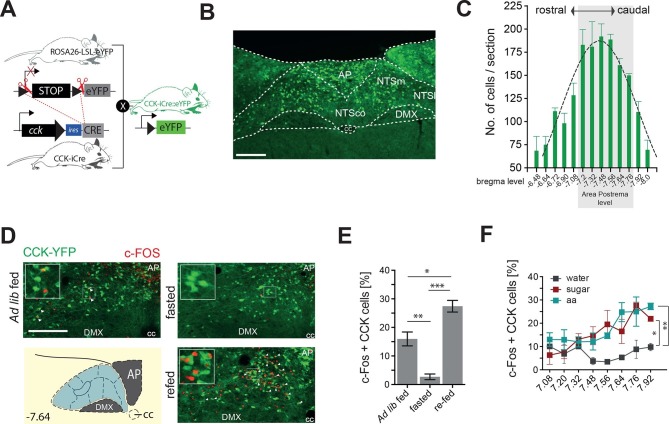
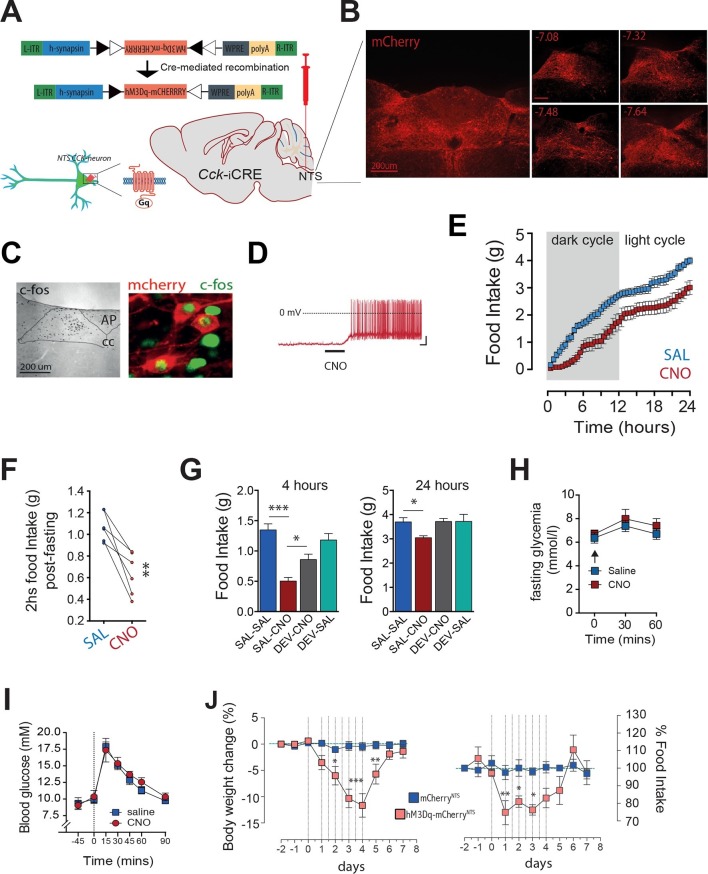
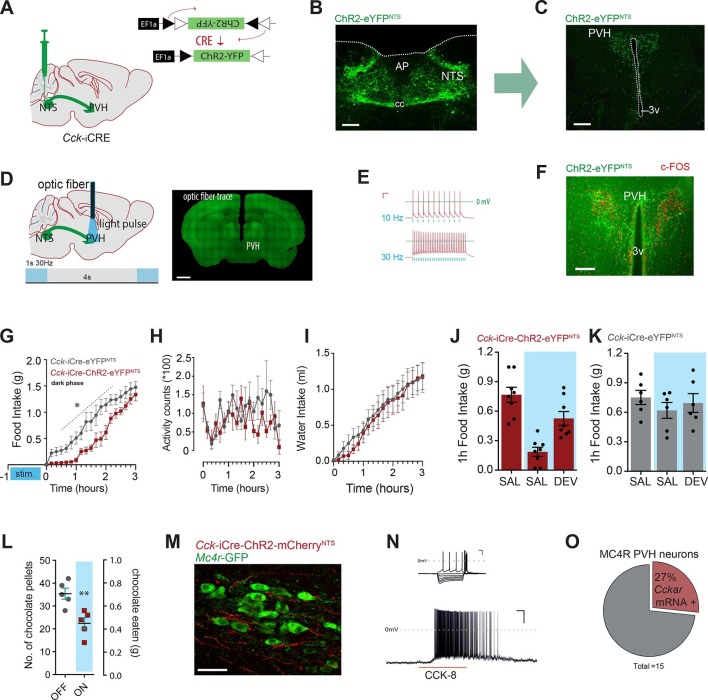
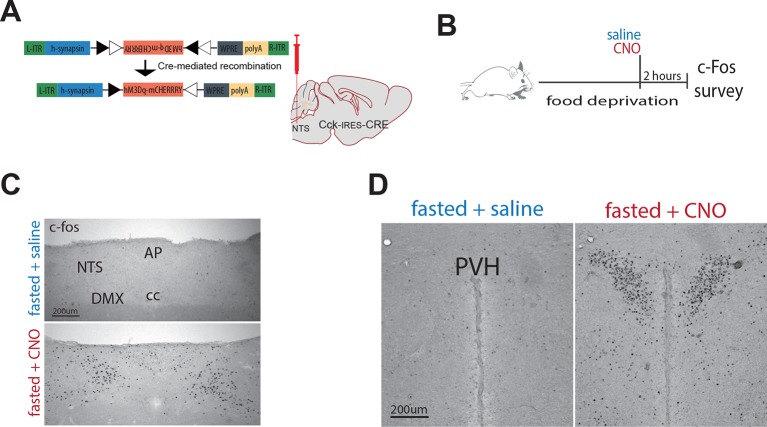
The nucleus of the solitary tract (NTS) is a key gateway for meal-related signals entering the brain from the periphery. However, the chemical mediators crucial to this process have not been fully elucidated. We reveal that a subset of NTS neurons containing cholecystokinin (CCK(NTS)) is responsive to nutritional state and that their activation reduces appetite and body weight in mice. Cell-specific anterograde tracing revealed that CCK(NTS) neurons provide a distinctive innervation of the paraventricular nucleus of the hypothalamus (PVH), with fibers and varicosities in close apposition to a subset of melanocortin-4 receptor (MC4R(PVH)) cells, which are also responsive to CCK. Optogenetic activation of CCK(NTS) axon terminals within the PVH reveal the satiating function of CCK(NTS) neurons to be mediated by a CCK(NTS)→PVH pathway that also encodes positive valence. These data identify the functional significance of CCK(NTS) neurons and reveal a sufficient and discrete NTS to hypothalamus circuit controlling appetite.
Keywords: feeding behavior; mouse; neuronal circuits; neuropeptides; neuroscience.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
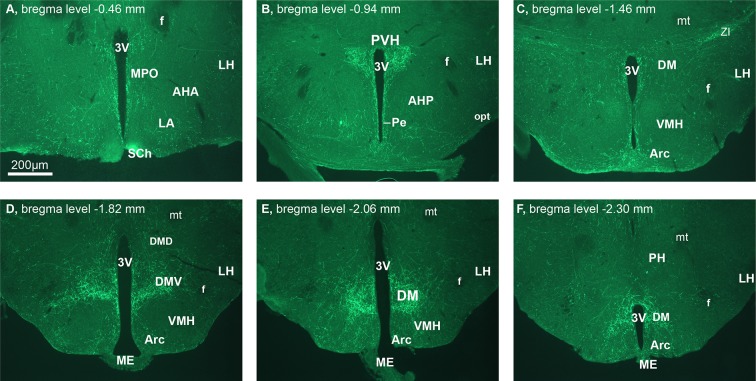
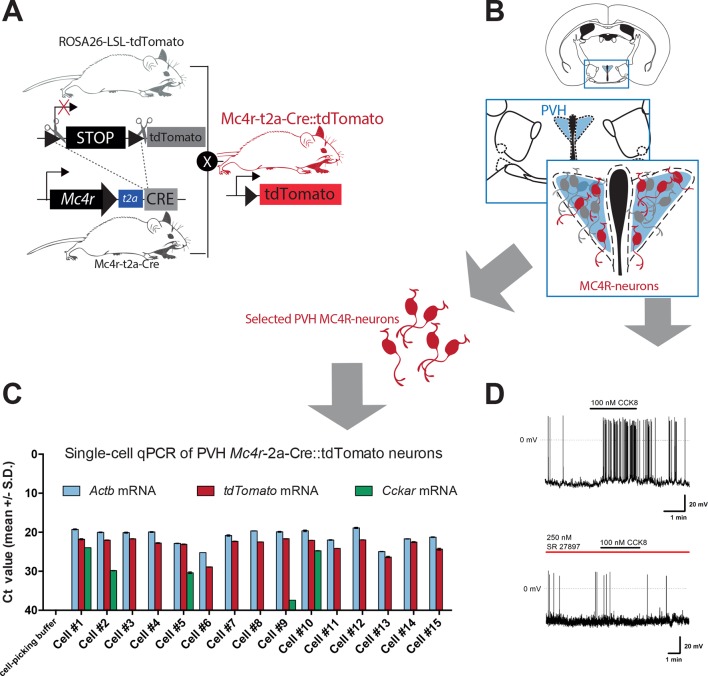
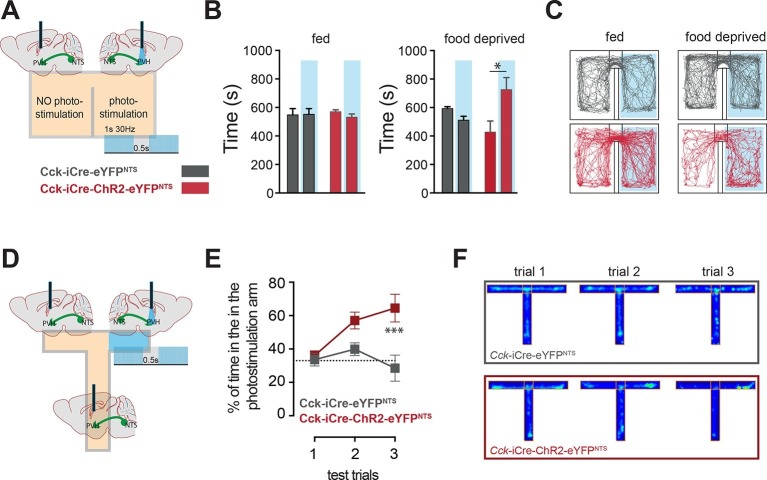
Comment in
-
Decoding obesity in the brainstem.Elife. 2016 May 9;5:e16393. doi: 10.7554/eLife.16393. Elife. 2016. PMID: 27156646 Free PMC article.
References
-
- Alexander GM, Rogan SC, Abbas AI, Armbruster BN, Pei Y, Allen JA, Nonneman RJ, Hartmann J, Moy SS, Nicolelis MA, McNamara JO, Roth BL. Remote control of neuronal activity in transgenic mice expressing evolved G protein-coupled receptors. Neuron. 2009;63:27–39. doi: 10.1016/j.neuron.2009.06.014. - DOI - PMC - PubMed
-
- Appleyard SM, Bailey TW, Doyle MW, Jin YH, Smart JL, Low MJ, Andresen MC. Proopiomelanocortin neurons in nucleus tractus solitarius are activated by visceral afferents: regulation by cholecystokinin and opioids. Journal of Neuroscience. 2005;25:3578–3585. doi: 10.1523/JNEUROSCI.4177-04.2005. - DOI - PMC - PubMed
-
- Balthasar N, Dalgaard LT, Lee CE, Yu J, Funahashi H, Williams T, Ferreira M, Tang V, McGovern RA, Kenny CD, Christiansen LM, Edelstein E, Choi B, Boss O, Aschkenasi C, Zhang Chen-yu, Mountjoy K, Kishi T, Elmquist JK, Lowell BB. Divergence of melanocortin pathways in the control of food intake and energy expenditure. Cell. 2005;123:493–505. doi: 10.1016/j.cell.2005.08.035. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
