

Disulfide Cross-linking of a Multidrug and Toxic Compound Extrusion Transporter Impacts Multidrug Efflux
- PMID: 26975373
- PMCID: PMC4850317
- DOI: 10.1074/jbc.M116.715227
Disulfide Cross-linking of a Multidrug and Toxic Compound Extrusion Transporter Impacts Multidrug Efflux
Abstract
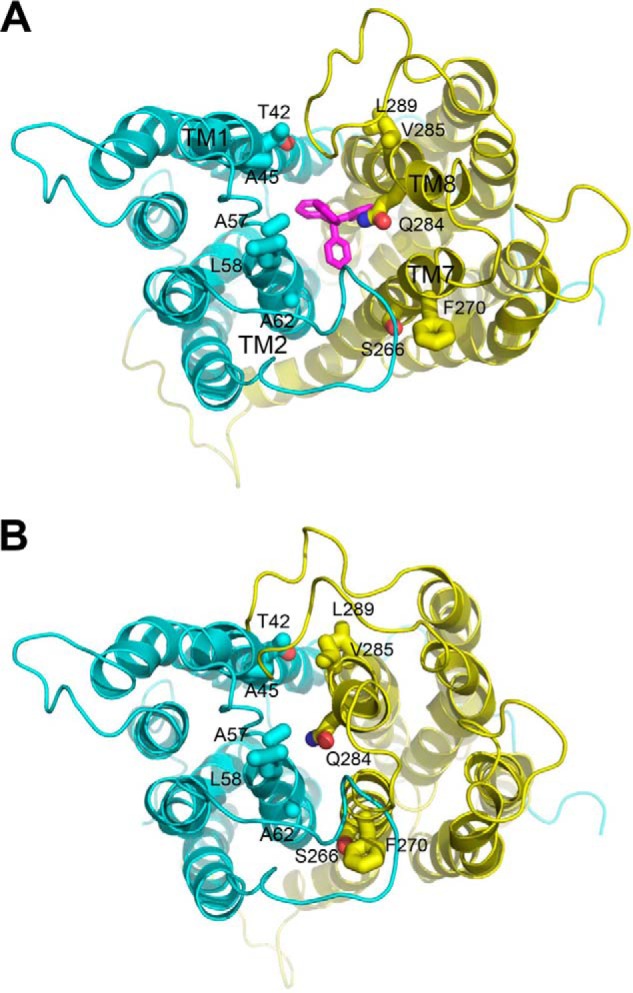
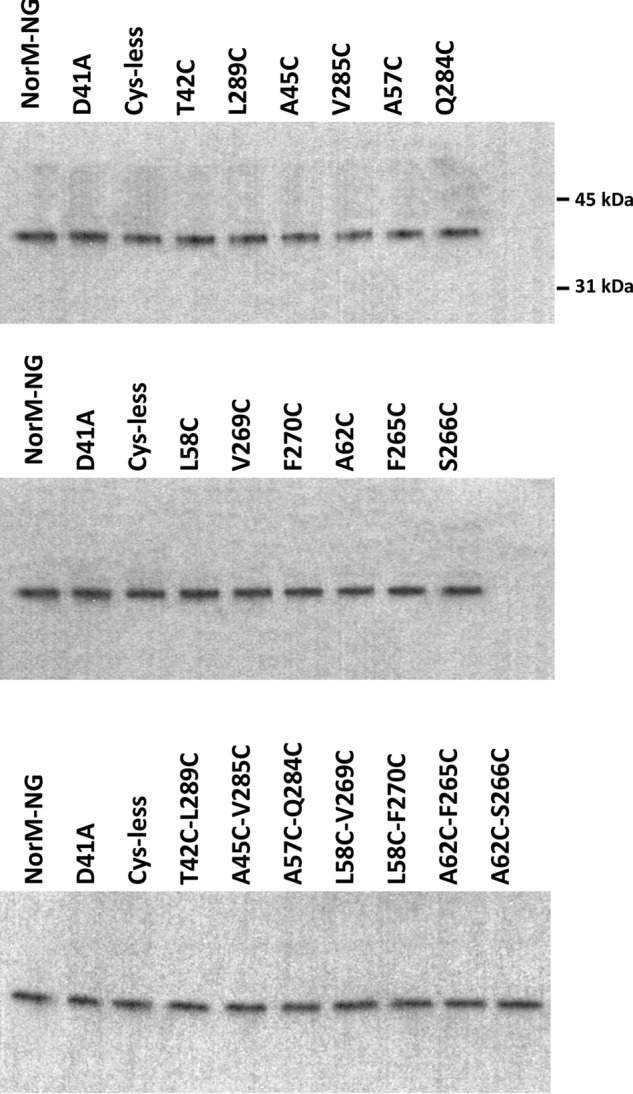
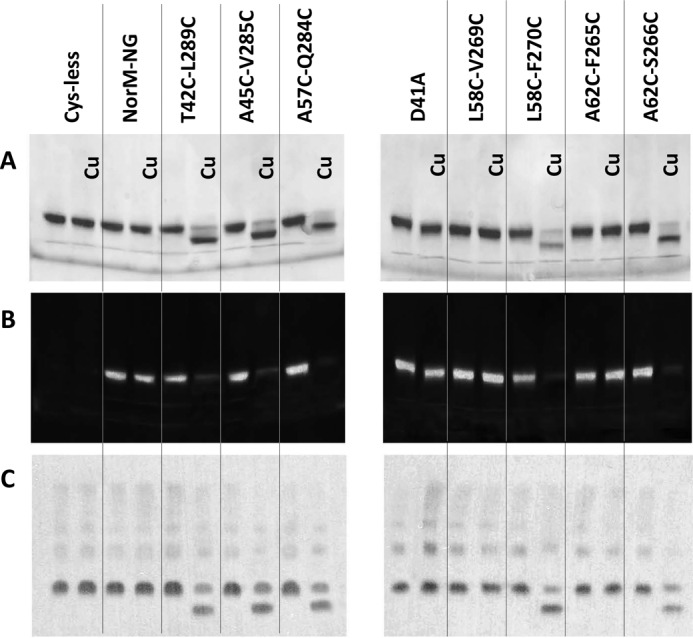
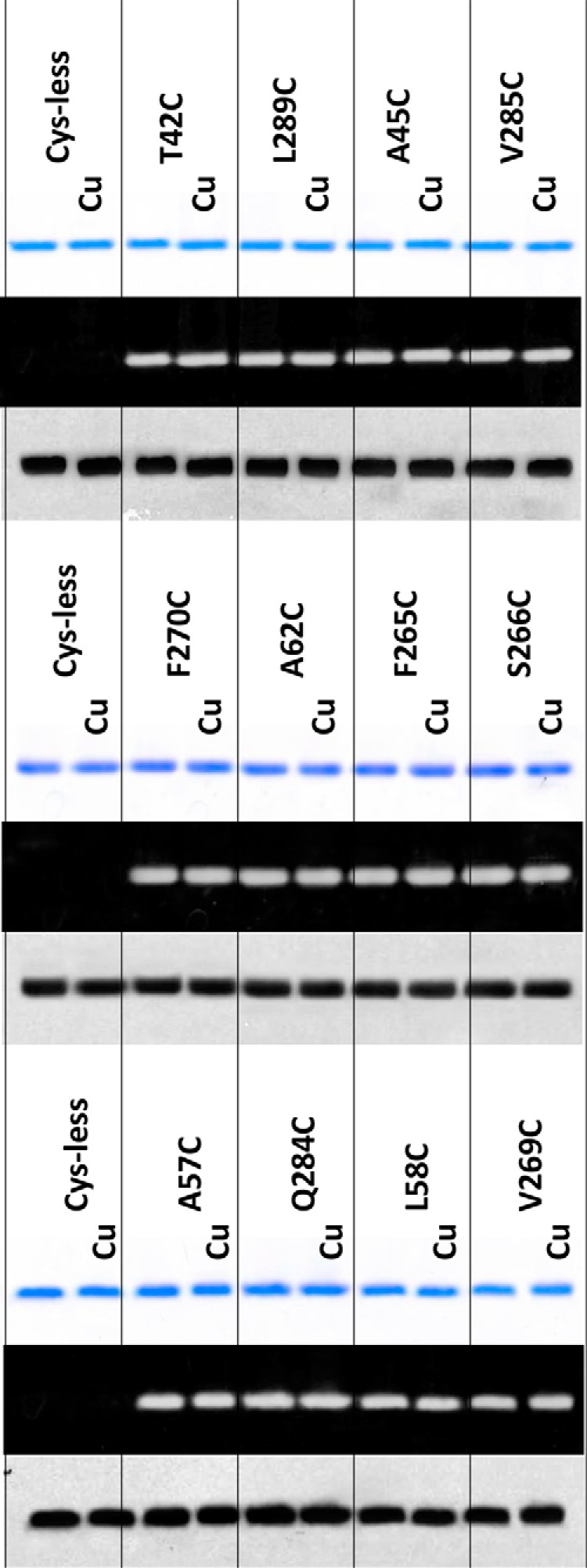
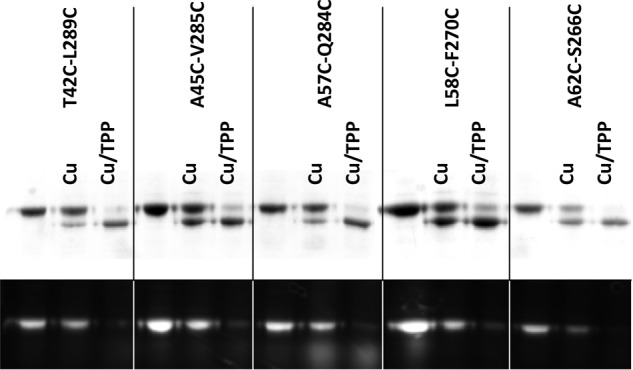
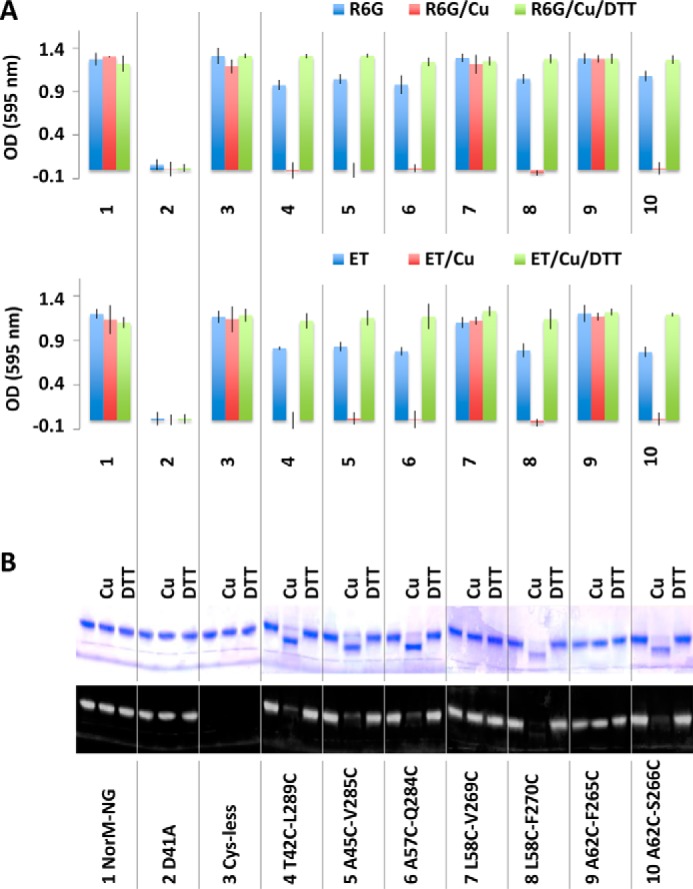
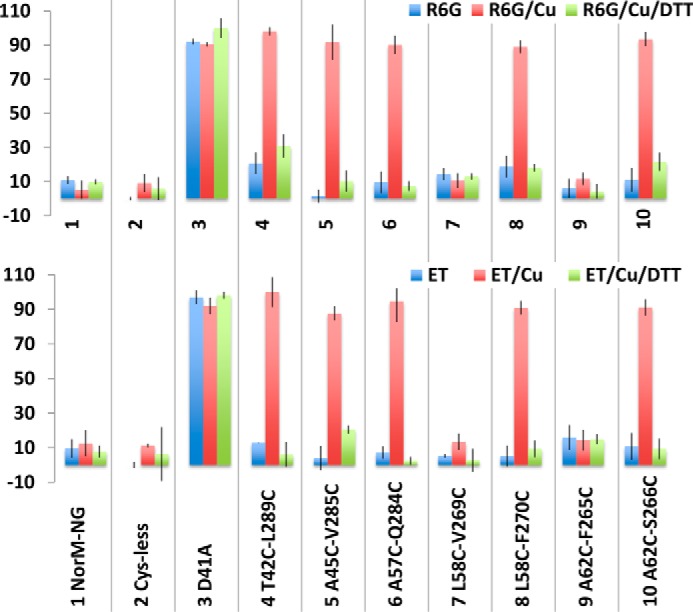
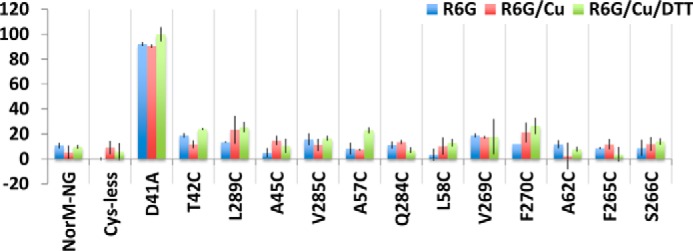
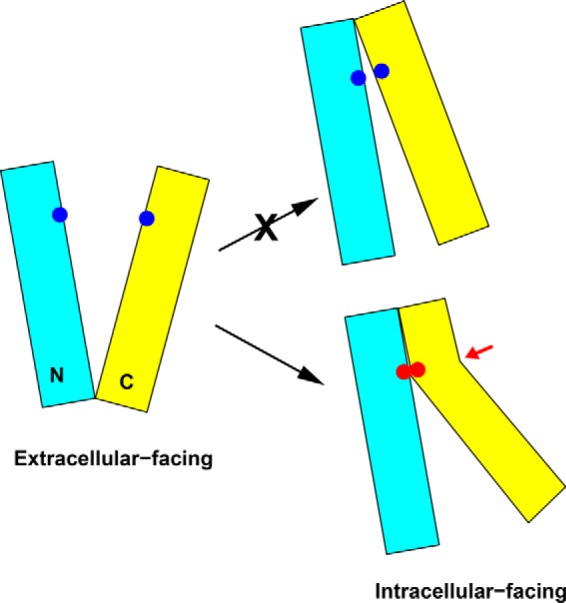
Multidrug and toxic compound extrusion (MATE) transporters contribute to multidrug resistance by extruding different drugs across cell membranes. The MATE transporters alternate between their extracellular and intracellular facing conformations to propel drug export, but how these structural changes occur is unclear. Here we combine site-specific cross-linking and functional studies to probe the movement of transmembrane helices in NorM from Neiserria gonorrheae (NorM-NG), a MATE transporter with known extracellular facing structure. We generated an active, cysteine-less NorM-NG and conducted pairwise cysteine mutagenesis on this variant. We found that copper phenanthroline catalyzed disulfide bond formation within five cysteine pairs and increased the electrophoretic mobility of the corresponding mutants. Furthermore, copper phenanthroline abolished the activity of the five paired cysteine mutants, suggesting that these substituted amino acids come in spatial proximity during transport, and the proximity changes are functionally indispensable. Our data also implied that the substrate-binding transmembrane helices move up to 10 Å in NorM-NG during transport and afforded distance restraints for modeling the intracellular facing transporter, thereby casting new light on the underlying mechanism.
Keywords: conformational change; cysteine cross-linking; drug resistance; exchanger; membrane protein; multidrug efflux; multidrug transporter.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Higgins C. F. (2007) Multiple molecular mechanisms for multidrug resistance transporters. Nature 446, 749–757 - PubMed
-
- Brown M. H., Paulsen I. T., and Skurray R. A. (1999) The multidrug efflux protein NorM is a prototype of a new family of transporters. Mol. Microbiol. 31, 394–395 - PubMed
-
- Omote H., Hiasa M., Matsumoto T., Otsuka M., and Moriyama Y. (2006) The MATE proteins as fundamental transporters of metabolic and xenobiotic organic cations. Trends Pharmacol. Sci. 27, 587–593 - PubMed
-
- Kuroda T., and Tsuchiya T. (2009) Multidrug efflux transporters in the MATE family. Biochim. Biophys. Acta 1794, 763–768 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
