

Differential Arc expression in the hippocampus and striatum during the transition from attentive to automatic navigation on a plus maze
- PMID: 26976088
- PMCID: PMC4883580
- DOI: 10.1016/j.nlm.2016.03.008
Differential Arc expression in the hippocampus and striatum during the transition from attentive to automatic navigation on a plus maze
Erratum in
-
Corrigendum to "Differential Arc expression in the hippocampus and striatum during the transition from attentive to automatic navigation on a plus maze" [Neurobiol. Learn. Mem. 131 (2016) 36-45].Neurobiol Learn Mem. 2016 Jul;132:67. doi: 10.1016/j.nlm.2016.05.013. Epub 2016 Jun 4. Neurobiol Learn Mem. 2016. PMID: 27267569 No abstract available.
Abstract
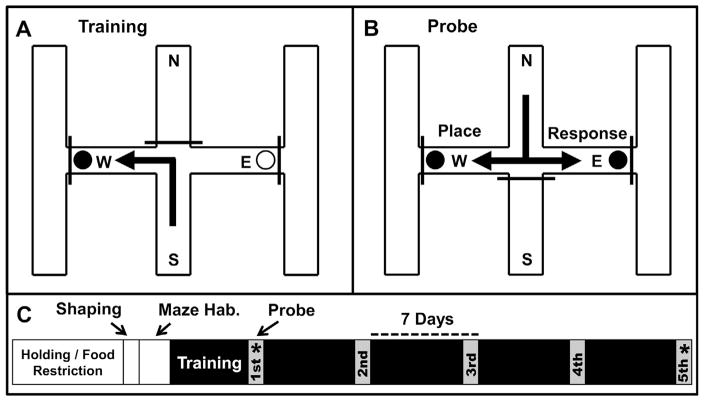
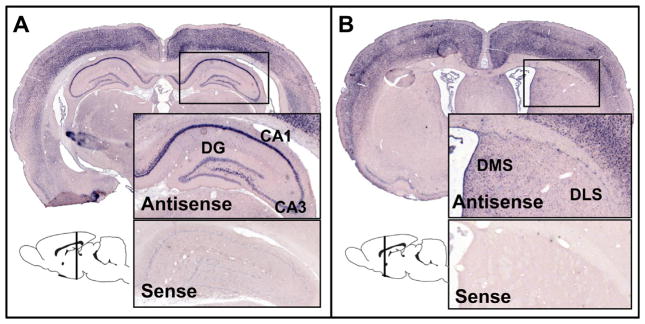
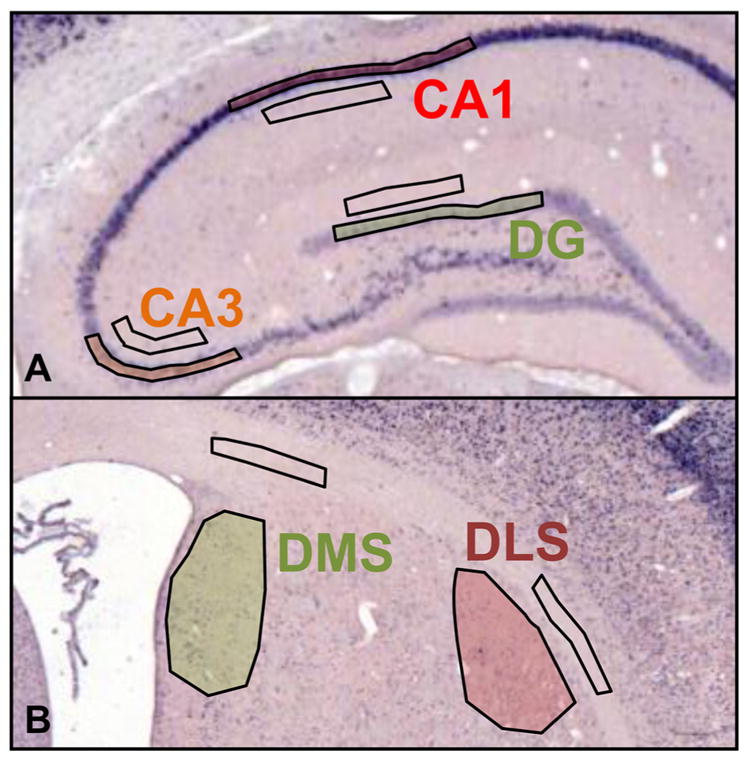
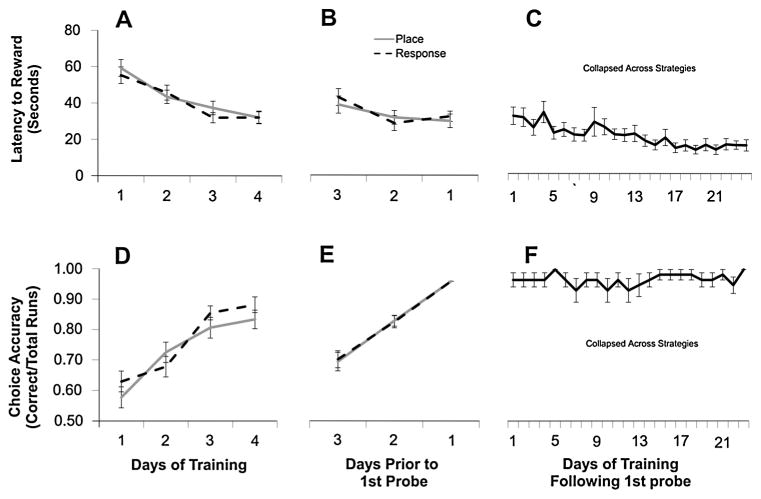
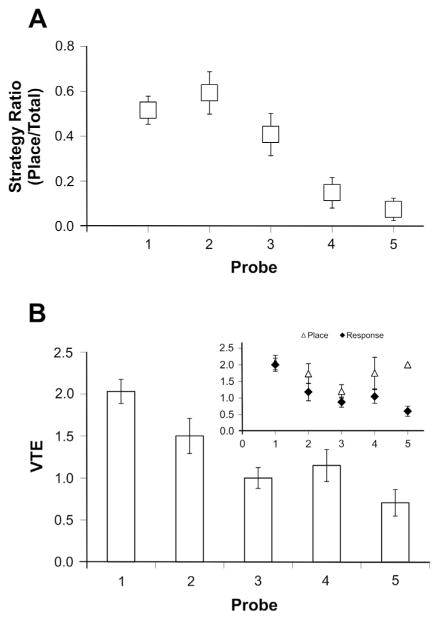
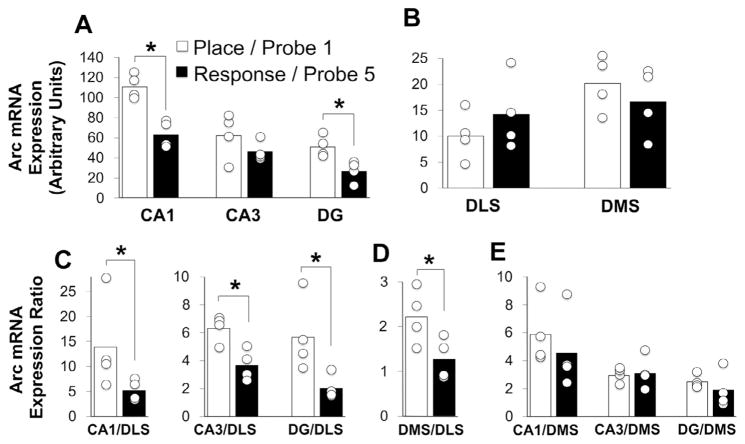
The strategies utilized to effectively perform a given task change with practice and experience. During a spatial navigation task, with relatively little training, performance is typically attentive enabling an individual to locate the position of a goal by relying on spatial landmarks. These (place) strategies require an intact hippocampus. With task repetition, performance becomes automatic; the same goal is reached using a fixed response or sequence of actions. These (response) strategies require an intact striatum. The current work aims to understand the activation patterns across these neural structures during this experience-dependent strategy transition. This was accomplished by region-specific measurement of activity-dependent immediate early gene expression among rats trained to different degrees on a dual-solution task (i.e., a task that can be solved using either place or response navigation). As expected, rats increased their reliance on response navigation with extended task experience. In addition, dorsal hippocampal expression of the immediate early gene Arc was considerably reduced in rats that used a response strategy late in training (as compared with hippocampal expression in rats that used a place strategy early in training). In line with these data, vicarious trial and error, a behavior linked to hippocampal function, also decreased with task repetition. Although Arc mRNA expression in dorsal medial or lateral striatum alone did not correlate with training stage, the ratio of expression in the medial striatum to that in the lateral striatum was relatively high among rats that used a place strategy early in training as compared with the ratio among over-trained response rats. Altogether, these results identify specific changes in the activation of dissociated neural systems that may underlie the experience-dependent emergence of response-based automatic navigation.
Keywords: Activity-regulated gene expression; Hippocampus; Place and response navigation; Spatial memory; Striatum; Vicarious trial and error.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Immediate early gene activation in hippocampus and dorsal striatum: effects of explicit place and response training.Neurobiol Learn Mem. 2007 May;87(4):583-96. doi: 10.1016/j.nlm.2006.12.011. Epub 2007 Feb 21. Neurobiol Learn Mem. 2007. PMID: 17317230
-
Shifting between response and place strategies in maze navigation: Effects of training, cue availability and functional inactivation of striatum or hippocampus in rats.Neurobiol Learn Mem. 2020 Jan;167:107131. doi: 10.1016/j.nlm.2019.107131. Epub 2019 Nov 26. Neurobiol Learn Mem. 2020. PMID: 31783128
-
Differential induction of c-Jun and Fos-like proteins in rat hippocampus and dorsal striatum after training in two water maze tasks.Neurobiol Learn Mem. 2005 Sep;84(2):75-84. doi: 10.1016/j.nlm.2005.03.006. Neurobiol Learn Mem. 2005. PMID: 15936959
-
Parallel processing across neural systems: implications for a multiple memory system hypothesis.Neurobiol Learn Mem. 2004 Nov;82(3):278-98. doi: 10.1016/j.nlm.2004.07.007. Neurobiol Learn Mem. 2004. PMID: 15464410 Review.
-
Emergence of the Hippocampus as a Vector for Goal-Directed Spatial Navigation.Adv Neurobiol. 2024;41:39-61. doi: 10.1007/978-3-031-69188-1_2. Adv Neurobiol. 2024. PMID: 39589709 Review.
Cited by
-
Inactivation of the striatum in aged rats rescues their ability to learn a hippocampus-sensitive spatial navigation task.Neurobiol Learn Mem. 2020 Jul;172:107231. doi: 10.1016/j.nlm.2020.107231. Epub 2020 Apr 17. Neurobiol Learn Mem. 2020. PMID: 32305514 Free PMC article.
-
Involvement of lactate transport in two object recognition tasks that require either the hippocampus or striatum.Behav Neurosci. 2019 Apr;133(2):176-187. doi: 10.1037/bne0000304. Behav Neurosci. 2019. PMID: 30907617 Free PMC article.
-
Memory and eating: A bidirectional relationship implicated in obesity.Neurosci Biobehav Rev. 2022 Jan;132:110-129. doi: 10.1016/j.neubiorev.2021.10.051. Epub 2021 Nov 20. Neurosci Biobehav Rev. 2022. PMID: 34813827 Free PMC article. Review.
-
Arc/Arg3.1 mediates a critical period for spatial learning and hippocampal networks.Proc Natl Acad Sci U S A. 2018 Dec 4;115(49):12531-12536. doi: 10.1073/pnas.1810125115. Epub 2018 Nov 15. Proc Natl Acad Sci U S A. 2018. PMID: 30442670 Free PMC article.
-
Hippocampus-sensitive and striatum-sensitive learning one month after morphine or cocaine exposure in male rats.Pharmacol Biochem Behav. 2022 Jun;217:173392. doi: 10.1016/j.pbb.2022.173392. Epub 2022 May 2. Pharmacol Biochem Behav. 2022. PMID: 35513118 Free PMC article.
References
-
- Barnes CA, Suster MS, Shen J, McNaughton BL. Multistability of cognitive maps in the hippocampus of old rats. Nature. 1997;388:272–275. http://dx.doi.org/10.1038/40859. - DOI - PubMed
-
- Benjamini Y, Hochberg Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society Series B (Methodological) 1995;57:289–300. http://dx.doi.org/10.2307/2346101. - DOI
-
- Bramham CR, Alme MN, Bittins M, Kuipers SD, Nair RR, Pai B, et al. The Arc of synaptic memory. Experimental Brain Research. 2010;200:125–140. http://dx.doi.org/10.1007/s00221-009-1959-2. - DOI - PMC - PubMed
-
- Daberkow DP, Riedy MD, Kesner RP, Keefe KA. Arc mRNA induction in striatal efferent neurons associated with response learning. European Journal of Neuroscience. 2007;26:228–241. http://dx.doi.org/10.1111/j.1460-9568.2007.05630.x. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
