

Diverse high-torque bacterial flagellar motors assemble wider stator rings using a conserved protein scaffold
- PMID: 26976588
- PMCID: PMC4822576
- DOI: 10.1073/pnas.1518952113
Diverse high-torque bacterial flagellar motors assemble wider stator rings using a conserved protein scaffold
Erratum in
-
Correction for Beeby et al., Diverse high-torque bacterial flagellar motors assemble wider stator rings using a conserved protein scaffold.Proc Natl Acad Sci U S A. 2016 May 10;113(19):E2759. doi: 10.1073/pnas.1605597113. Epub 2016 May 2. Proc Natl Acad Sci U S A. 2016. PMID: 27140639 Free PMC article. No abstract available.
Abstract
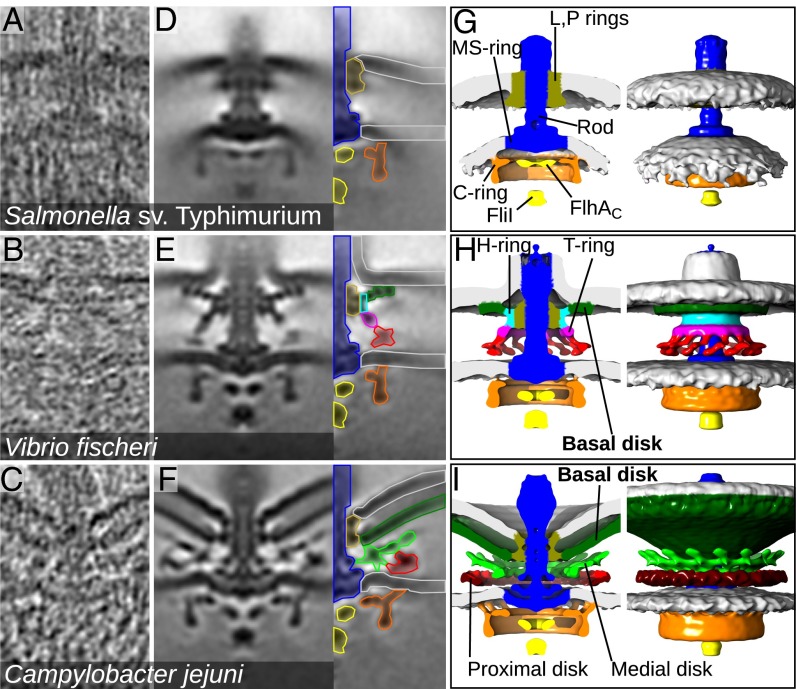
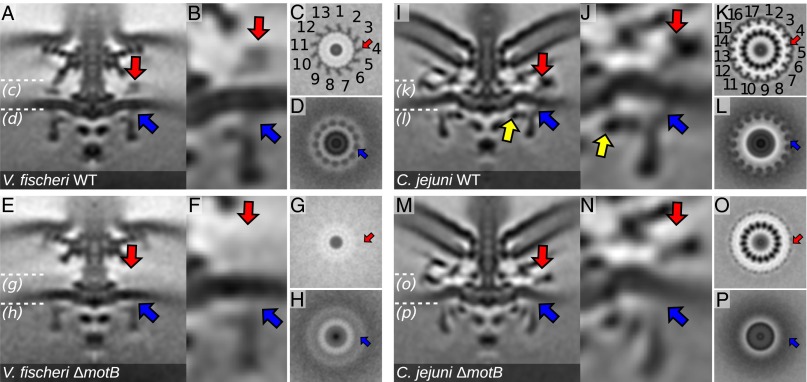
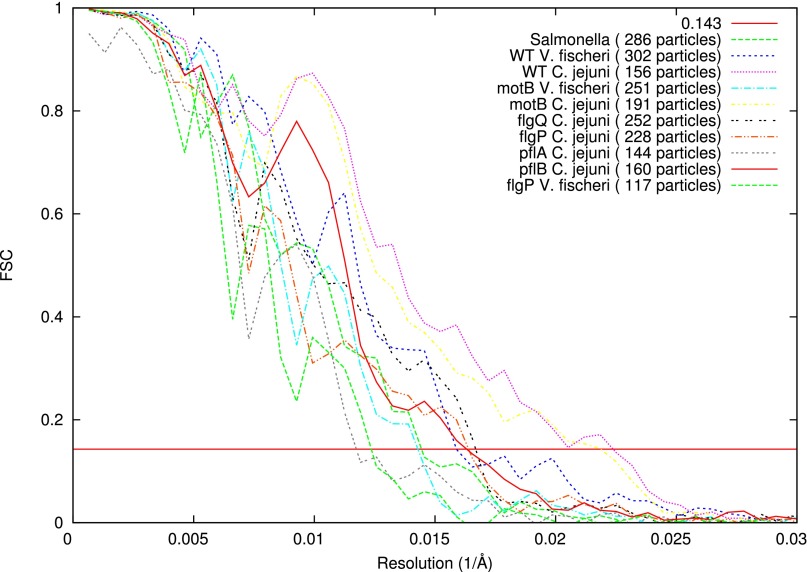
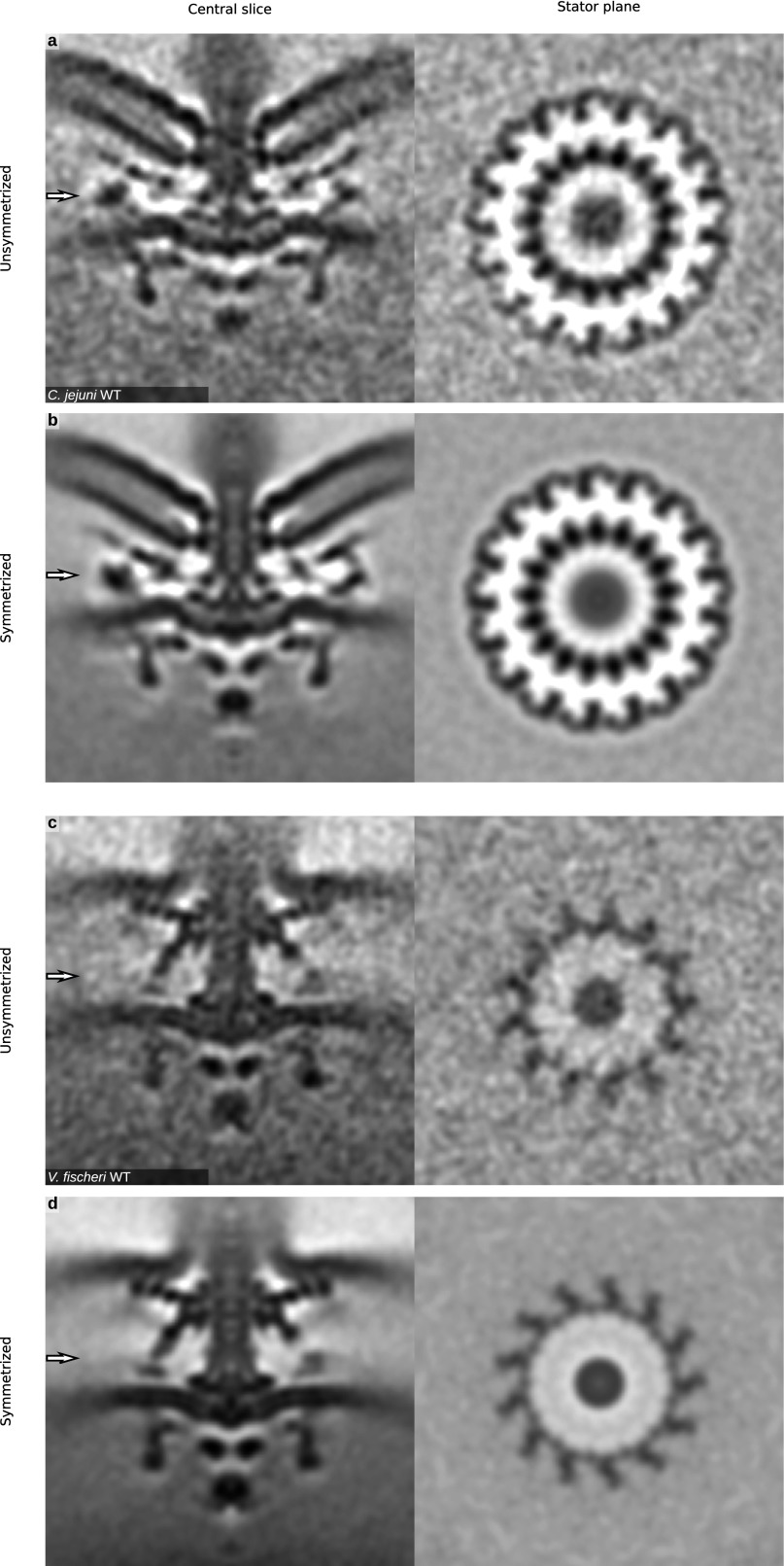
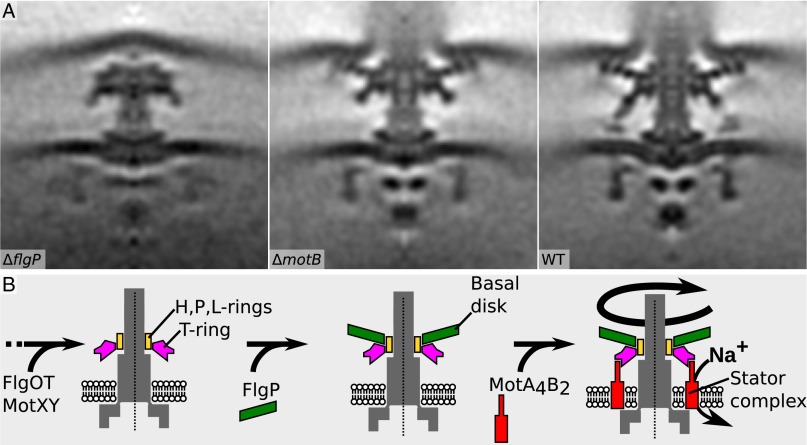
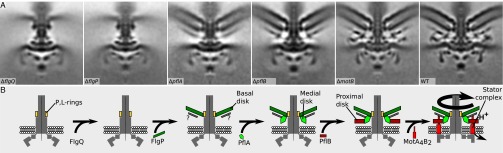
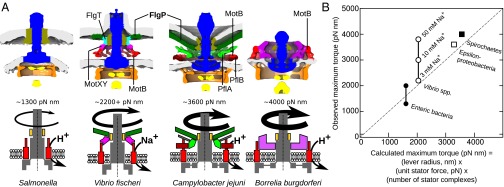
Although it is known that diverse bacterial flagellar motors produce different torques, the mechanism underlying torque variation is unknown. To understand this difference better, we combined genetic analyses with electron cryo-tomography subtomogram averaging to determine in situ structures of flagellar motors that produce different torques, from Campylobacter and Vibrio species. For the first time, to our knowledge, our results unambiguously locate the torque-generating stator complexes and show that diverse high-torque motors use variants of an ancestrally related family of structures to scaffold incorporation of additional stator complexes at wider radii from the axial driveshaft than in the model enteric motor. We identify the protein components of these additional scaffold structures and elucidate their sequential assembly, demonstrating that they are required for stator-complex incorporation. These proteins are widespread, suggesting that different bacteria have tailored torques to specific environments by scaffolding alternative stator placement and number. Our results quantitatively account for different motor torques, complete the assignment of the locations of the major flagellar components, and provide crucial constraints for understanding mechanisms of torque generation and the evolution of multiprotein complexes.
Keywords: Campylobacter; bacterial flagellar motors; electron cryo-tomography; macromolecular evolution; torque.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Comment in
-
Structural biology: ECT joins the rotary club.Nat Rev Microbiol. 2016 Apr;14(5):265. doi: 10.1038/nrmicro.2016.51. Epub 2016 Apr 12. Nat Rev Microbiol. 2016. PMID: 27067403 No abstract available.
References
-
- Li N, Kojima S, Homma M. Sodium-driven motor of the polar flagellum in marine bacteria Vibrio. Genes to Cells. 2011;16:985–999. - PubMed
-
- Ferrero RL, Lee A. Motility of Campylobacter jejuni in a viscous environment: Comparison with conventional rod-shaped bacteria. J Gen Microbiol. 1988;134(1):53–59. - PubMed
-
- Hazell SL, Lee A, Brady L, Hennessy W. Campylobacter pyloridis and gastritis: Association with intercellular spaces and adaptation to an environment of mucus as important factors in colonization of the gastric epithelium. J Infect Dis. 1986;153(4):658–663. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
