

Sex difference in physical activity, energy expenditure and obesity driven by a subpopulation of hypothalamic POMC neurons
- PMID: 26977396
- PMCID: PMC4770275
- DOI: 10.1016/j.molmet.2016.01.005
Sex difference in physical activity, energy expenditure and obesity driven by a subpopulation of hypothalamic POMC neurons
Abstract
Objective: Obesity is one of the primary healthcare challenges of the 21st century. Signals relaying information regarding energy needs are integrated within the brain to influence body weight. Central among these integration nodes are the brain pro-opiomelanocortin (POMC) peptides, perturbations of which disrupt energy balance and promote severe obesity. However, POMC neurons are neurochemically diverse and the crucial source of POMC peptides that regulate energy homeostasis and body weight remains to be fully clarified.
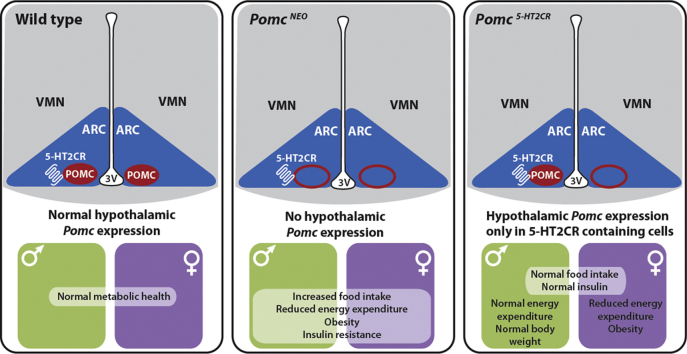
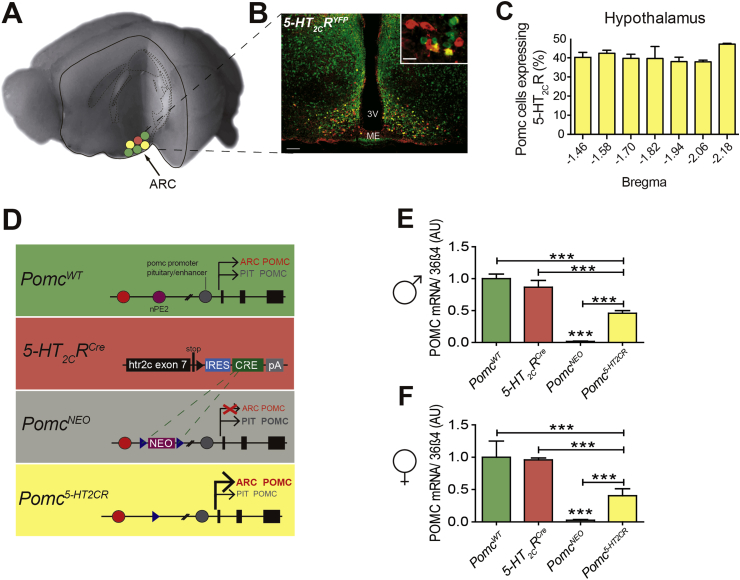
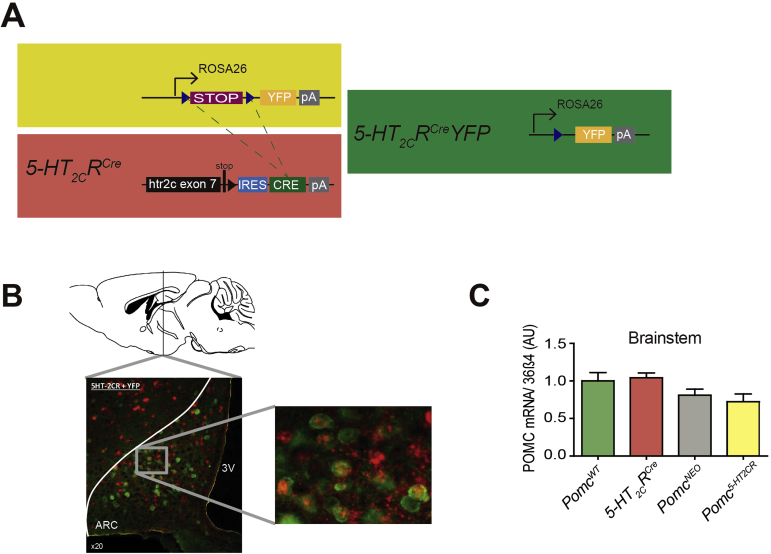
Methods: Given that a 5-hydroxytryptamine 2c receptor (5-HT2CR) agonist is a current obesity medication and 5-HT2CR agonist's effects on appetite are primarily mediated via POMC neurons, we hypothesized that a critical source of POMC regulating food intake and body weight is specifically synthesized in cells containing 5-HT2CRs. To exclusively manipulate Pomc synthesis only within 5-HT2CR containing cells, we generated a novel 5-HT 2C R (CRE) mouse line and intercrossed it with Cre recombinase-dependent and hypothalamic specific reactivatable Pomc (NEO) mice to restrict Pomc synthesis to the subset of hypothalamic cells containing 5-HT2CRs. This provided a means to clarify the specific contribution of a defined subgroup of POMC peptides in energy balance and body weight.
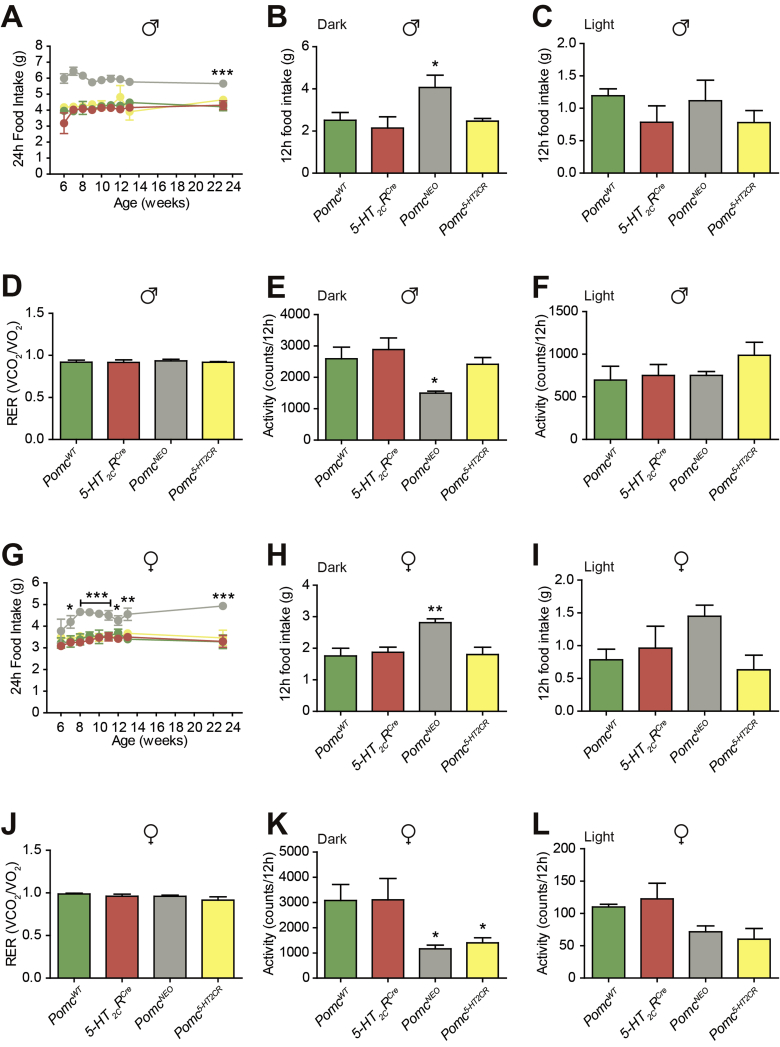
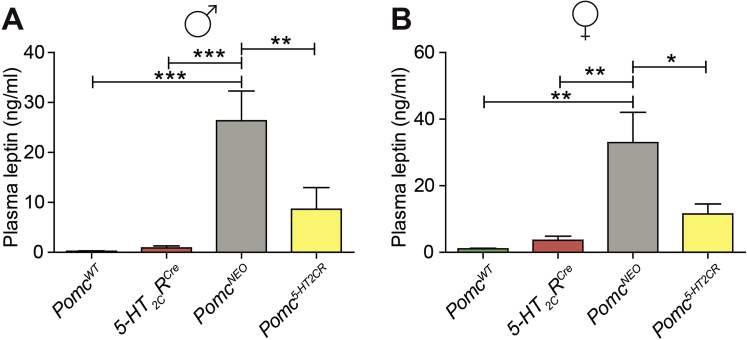
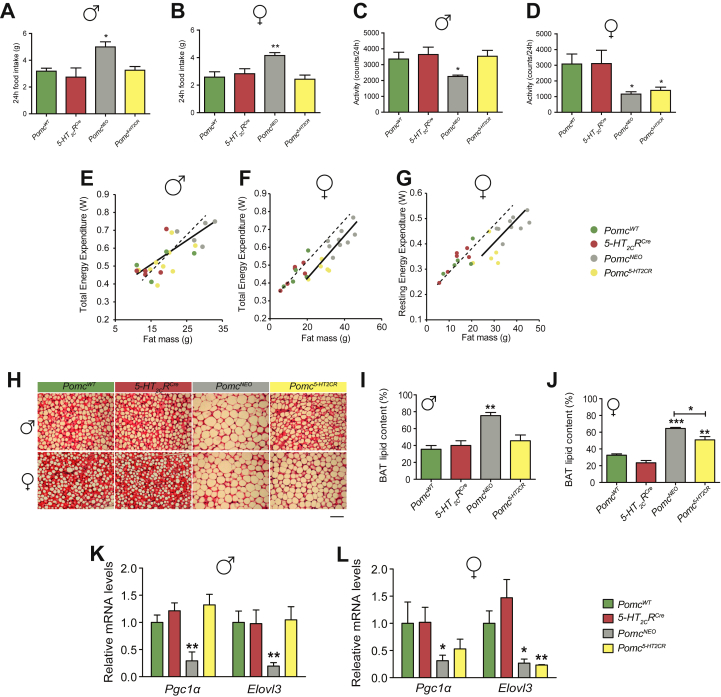
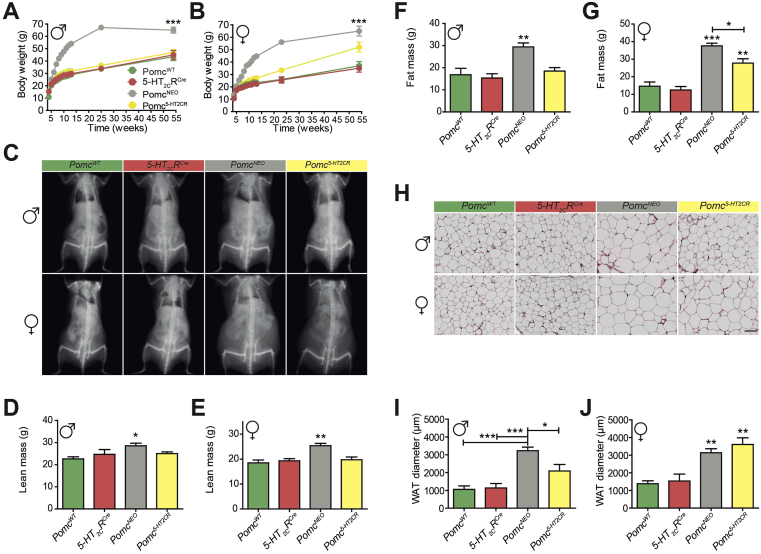
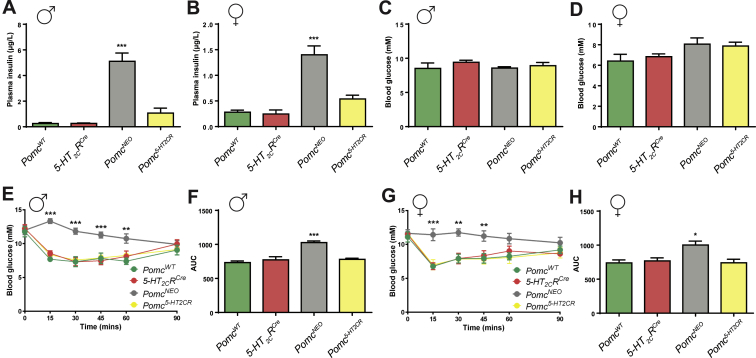
Results: Here we transform genetically programed obese and hyperinsulinemic male mice lacking hypothalamic Pomc with increased appetite, reduced physical activity and compromised brown adipose tissue (BAT) into lean, healthy mice via targeted restoration of Pomc function only within 5-HT2CR expressing cells. Remarkably, the same metabolic transformation does not occur in females, who despite corrected feeding behavior and normalized insulin levels remain physically inactive, have lower energy expenditure, compromised BAT and develop obesity.
Conclusions: These data provide support for the functional heterogeneity of hypothalamic POMC neurons, revealing that Pomc expression within 5-HT2CR expressing neurons is sufficient to regulate energy intake and insulin sensitivity in male and female mice. However, an unexpected sex difference in the function of this subset of POMC neurons was identified with regard to energy expenditure. We reveal that a large sex difference in physical activity, energy expenditure and the development of obesity is driven by this subpopulation, which constitutes approximately 40% of all POMC neurons in the hypothalamic arcuate nucleus. This may have broad implications for strategies utilized to combat obesity, which at present largely ignore the sex of the obese individual.
Keywords: 5-HT2c receptor; Brown adipose tissue; Energy expenditure; Hyperinsulinemia; Hypothalamus; Obesity; Pro-opiomelanocortin (Pomc); Sexual dimorphism.
Figures
References
-
- World Health Organisation . 2014. Global Health Observatory (GHO) obesity situation and trends.
-
- Yeo G.S., Heisler L.K. Unraveling the brain regulation of appetite: lessons from genetics. Nature Neuroscience. 2012 Oct;15(10):1343–1349. - PubMed
Grants and funding
- R01 DK066604/DK/NIDDK NIH HHS/United States
- 081713/WT_/Wellcome Trust/United Kingdom
- R01 DK056731/DK/NIDDK NIH HHS/United States
- R37 DK056731/DK/NIDDK NIH HHS/United States
- T32 NS076401/NS/NINDS NIH HHS/United States
- T32 GM008322/GM/NIGMS NIH HHS/United States
- P30 DK034933/DK/NIDDK NIH HHS/United States
- R56 DK066604/DK/NIDDK NIH HHS/United States
- R01 DK068400/DK/NIDDK NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- MC_UU_12012/5/MRC_/Medical Research Council/United Kingdom
- 098012/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
