

Review
doi: 10.1038/nrm.2016.24.
Epub 2016 Mar 16.
Adult cell plasticity in vivo: de-differentiation and transdifferentiation are back in style
Affiliations
- PMID: 26979497
- PMCID: PMC5818993
- DOI: 10.1038/nrm.2016.24
Item in Clipboard
Review
Adult cell plasticity in vivo: de-differentiation and transdifferentiation are back in style
Nat Rev Mol Cell Biol.
2016 Jul.
Abstract
Biologists have long been intrigued by the possibility that cells can change their identity, a phenomenon known as cellular plasticity. The discovery that terminally differentiated cells can be experimentally coaxed to become pluripotent has invigorated the field, and recent studies have demonstrated that changes in cell identity are not limited to the laboratory. Specifically, certain adult cells retain the capacity to de-differentiate or transdifferentiate under physiological conditions, as part of an organ's normal injury response. Recent studies have highlighted the extent to which cell plasticity contributes to tissue homeostasis, findings that have implications for cell-based therapy.
Figures
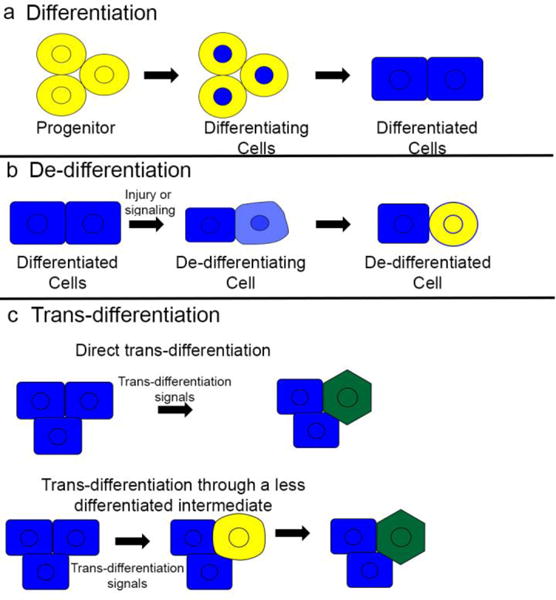
(a) During development and regeneration, progenitors and stem cells differentiate to form mature, differentiated cells. (b) In some cases of injury or stem cell ablation differentiated cells may de-differentiate, returning to an earlier fate and becoming progenitor cells again. (c) Injury and ablation may also induce differentiated cells to take on a different mature cell fate, a process known as trans-differentiation. Trans-differentiation may either occur directly, without any reversion to immature phenotypes, or through a de-differentiation step before cells re-differentiate to a new mature phenotype.
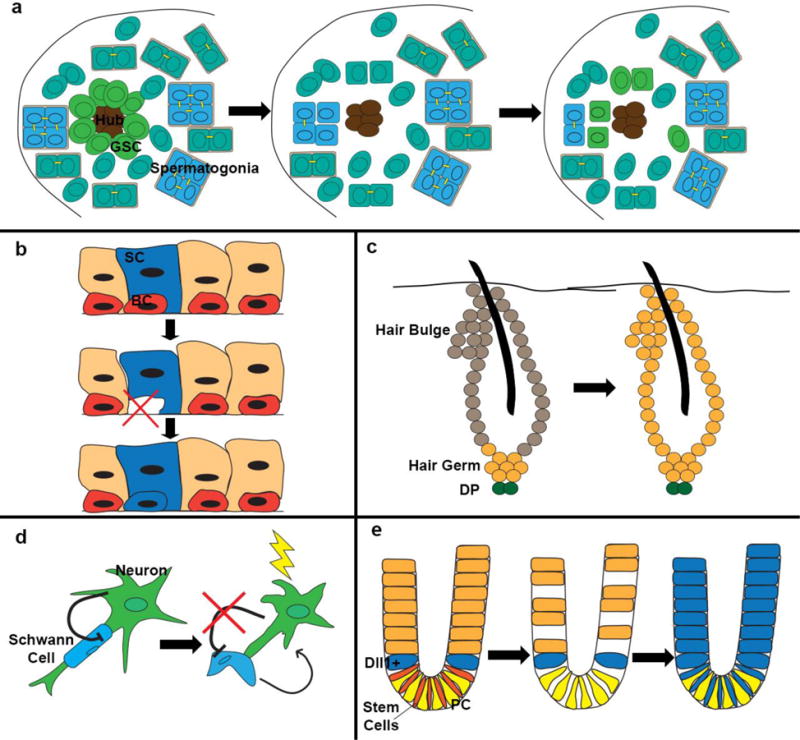
(a) Following loss of germ stem cells, differentiating spermatogonia may break apart from their multi-cell cysts to de-differentiate and form new germ stem cells around the hub. (b) In the lung airway, ablation of basal stem cells (BC) induces differentiated secretory cells (SC) to de-differentiate and form new BC. (c) During the hair cycle, hair germ cells give rise to transiently amplifying cells that make up the hair. Bulge cells contribute to the outer root sheath and to some cells of the hair germ for the next cycle. Following ablation of bulge cells, hair germ cells can de-differentiate and give rise to new bulge cells. DP: dermal papilla. (d) Following injury to the neuron, Schwann cells disassemble from the neuron and de-differentiate (a process normally inhibited by signals from the neuron), secreting factors to help with neuron regeneration. (e) In the intestine, radiation injury, which ablates crypt stem cells and cells further up in the crypt, exposes Dll1+ secretory cells to new signals from the paneth cells (PC) in the crypt which induce de-differentiation and expansion of the cell’s progeny up the crypt, as shown by lineage tracing.
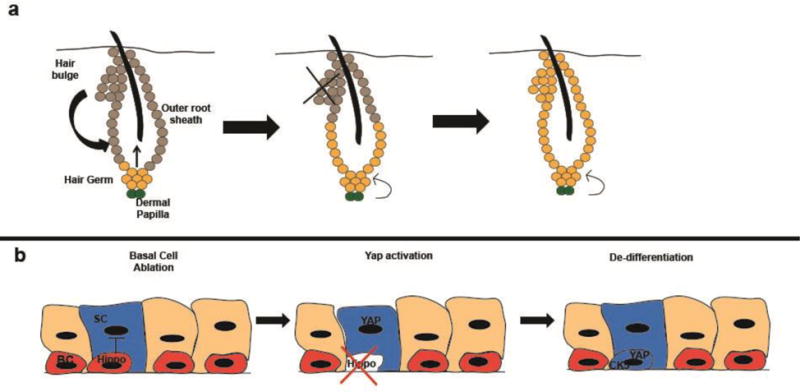
De-differentiation is induced as cells are exposed to new de-differentiation signals or have inhibitory signals silenced. (a), During the normal cycle of hair growth and regeneration, the hair germ produces transient amplifying cells that make up the hair while the hair bulge contributes to the outer root sheath. Following experimental ablation of the bulge, hair germ cells can de-differentiate and give rise to new bulge cells. This process relies on signals from the dermal papillae, as mechanical separation of the hair germ and dermal papilla prevents de-differentiation. (b), Basal cells adjacent to secretory cells (with one secretory cell genetically labeled in the diagram) secrete signals, possibly Hippo, preventing de-differentiation of secretory cells. Following ablation of basal cells, these signals are lost and secretory cell de-differentiation is no longer inhibited, allowing accumulation of the YAP transcription factor, expression of basal cell genes such as CK5, and de-differentiation of secretory cells into basal cells. These dedifferentiated basal cells can then differentiate to form new secretory cells. BC= basal cell, SC= secretory cell
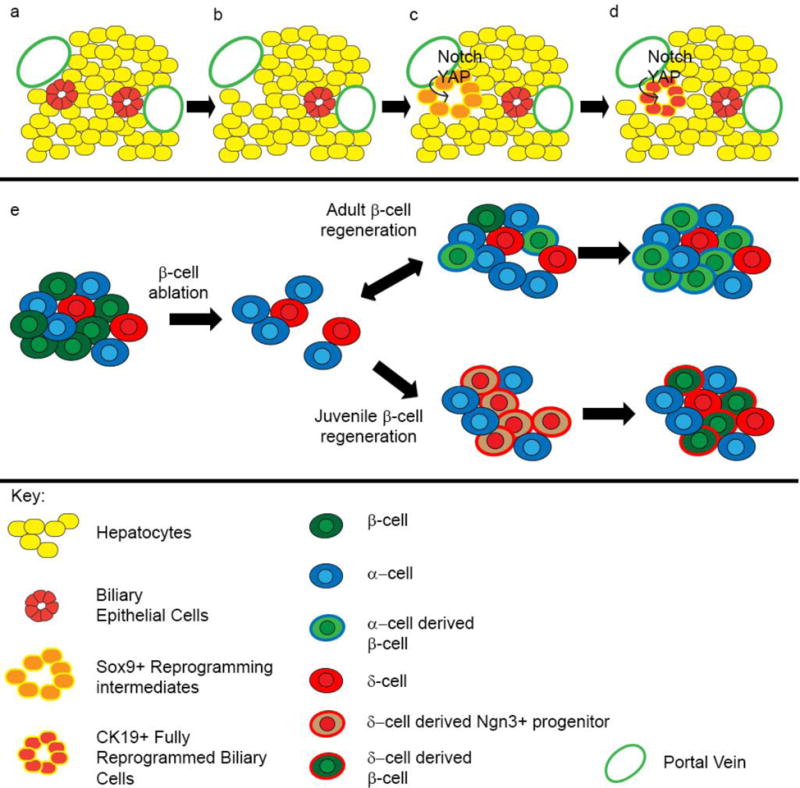
Hepatocytes trans-differentiate into biliary cells following liver injury. (a) Hepatic tissue prior to injury, with hepatocytes in yellow and biliary epithelial cells (bile ducts) in red. (b) Following injury and bile duct damage, (c) hepatocytes receive signals including Notch and activation of the YAP transcription factor that induce their trans-differentiation into biliary cells (shown in orange). During the trans-differentiation process, hepatocytes turn on a subset of “early” biliary genes, such as Sox9, which are specific to the new cell fate. (d) Over time trans-differentiated hepatocytes become mature bile ducts, expressing “late” bile duct markers such as CK19. (e) Normal pancreatic islets contain β, α, and δ cells. Following experimental ablation of β-cells, adult and juvenile islets follow different pathways to regenerate the missing tissue. In adult islets, α cells proliferate and begin to trans-differentiate directly in insulin-producing cells. Although these trans-differentiated cells are functional and produce insulin, they still maintain some α-gene expression, suggesting they are not fully trans-differentiated. In juvenile islets, δ cells proliferate and turn on δ cell-specific genes. These cells de-differentiate to Ngn3+ progenitor cells, then re-differentiate as β-cells. In this example, different pathways may lead to restoration of islet cells.
References
-
- Holliday R. Epigenetics: a historical overview. Epigenetics. 2006;1:76–80. - PubMed
-
- Gurdon JB. The developmental capacity of nuclei taken from intestinal epithelium cells of feeding tadpoles. J Embryol Exp Morphol. 1962;10:622–40. - PubMed
-
- Raff M. Adult stem cell plasticity: fact or artifact? Annu Rev Cell Dev Biol. 2003;19:1–22. - PubMed
-
- Jopling C, Boue S, Izpisua Belmonte JC. Dedifferentiation, transdifferentiation and reprogramming: three routes to regeneration. Nat Rev Mol Cell Biol. 2011;12:79–89. - PubMed
-
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–76. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
