

A phosphotyrosine switch regulates organic cation transporters
- PMID: 26979622
- PMCID: PMC4799362
- DOI: 10.1038/ncomms10880
A phosphotyrosine switch regulates organic cation transporters
Abstract
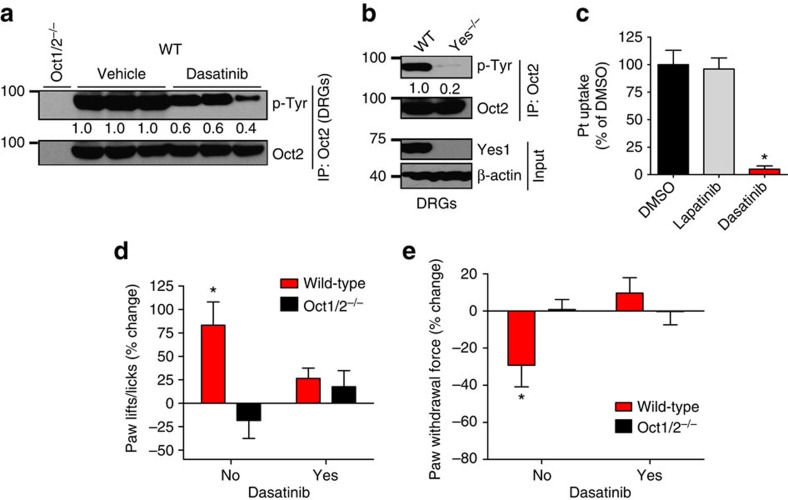
Membrane transporters are key determinants of therapeutic outcomes. They regulate systemic and cellular drug levels influencing efficacy as well as toxicities. Here we report a unique phosphorylation-dependent interaction between drug transporters and tyrosine kinase inhibitors (TKIs), which has uncovered widespread phosphotyrosine-mediated regulation of drug transporters. We initially found that organic cation transporters (OCTs), uptake carriers of metformin and oxaliplatin, were inhibited by several clinically used TKIs. Mechanistic studies showed that these TKIs inhibit the Src family kinase Yes1, which was found to be essential for OCT2 tyrosine phosphorylation and function. Yes1 inhibition in vivo diminished OCT2 activity, significantly mitigating oxaliplatin-induced acute sensory neuropathy. Along with OCT2, other SLC-family drug transporters are potentially part of an extensive 'transporter-phosphoproteome' with unique susceptibility to TKIs. On the basis of these findings we propose that TKIs, an important and rapidly expanding class of therapeutics, can functionally modulate pharmacologically important proteins by inhibiting protein kinases essential for their post-translational regulation.
Figures
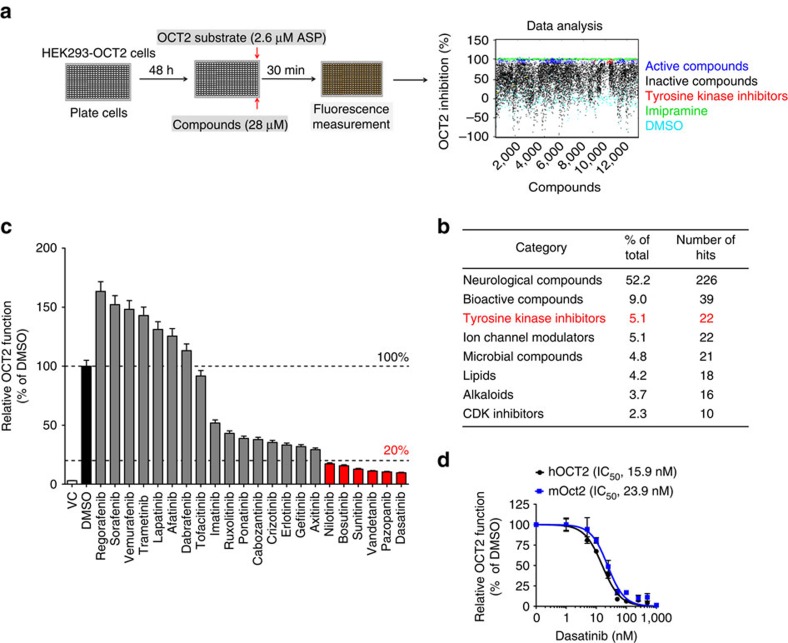
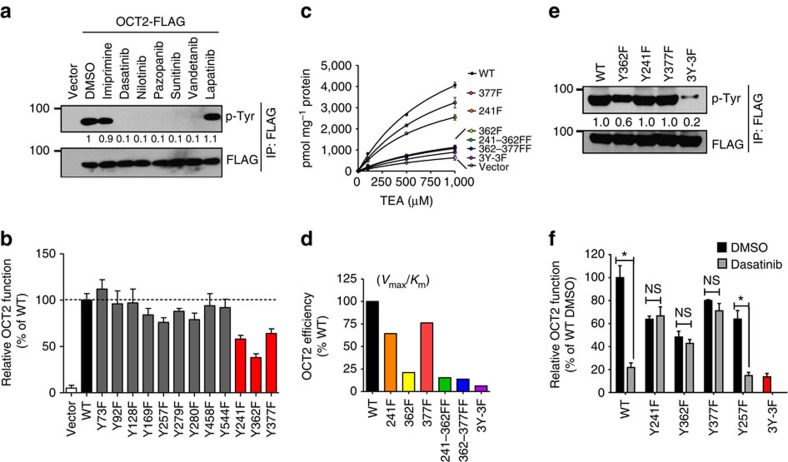
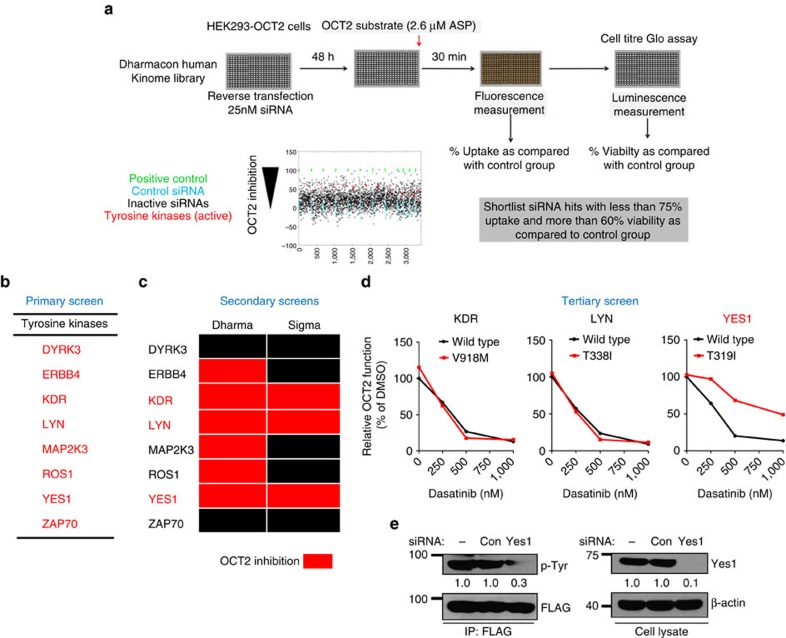

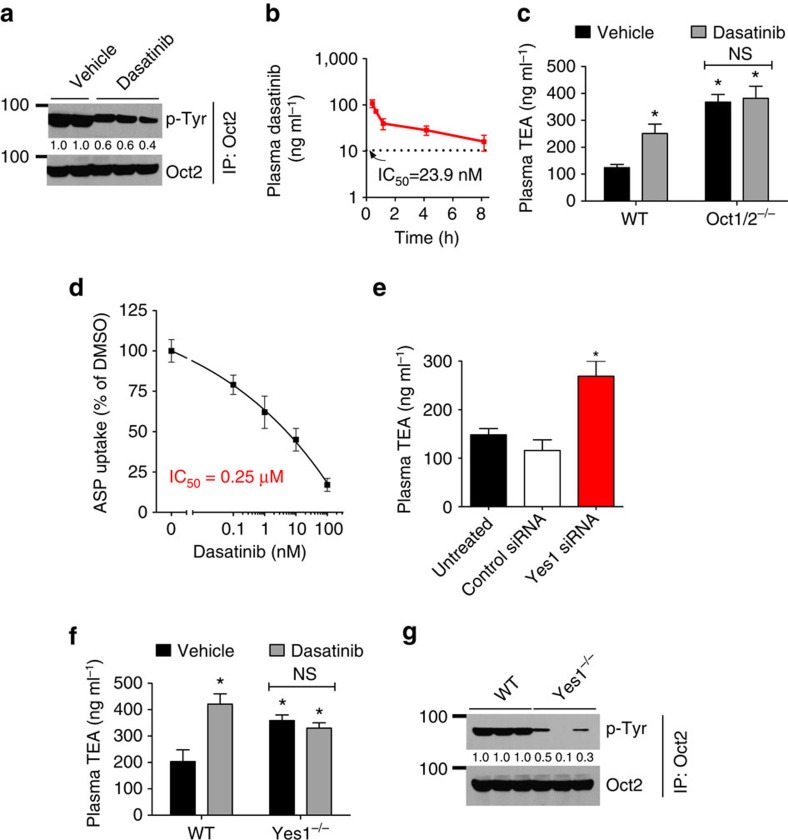
References
-
- Singer S. J. & Nicolson G. L. The fluid mosaic model of the structure of cell membranes. Science 175, 720–731 (1972). - PubMed
-
- Grecco H. E., Schmick M. & Bastiaens P. I. Signaling from the living plasma membrane. Cell 144, 897–909 (2011). - PubMed
-
- Kaback H. R., Sahin-Toth M. & Weinglass A. B. The kamikaze approach to membrane transport. Nat. Rev. Mol. Cell Biol. 2, 610–620 (2001). - PubMed
-
- Borst P. & Elferink R. O. Mammalian ABC transporters in health and disease. Annu. Rev. Biochem. 71, 537–592 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
