

C9orf72 ablation causes immune dysregulation characterized by leukocyte expansion, autoantibody production, and glomerulonephropathy in mice
- PMID: 26979938
- PMCID: PMC4793236
- DOI: 10.1038/srep23204
C9orf72 ablation causes immune dysregulation characterized by leukocyte expansion, autoantibody production, and glomerulonephropathy in mice
Abstract
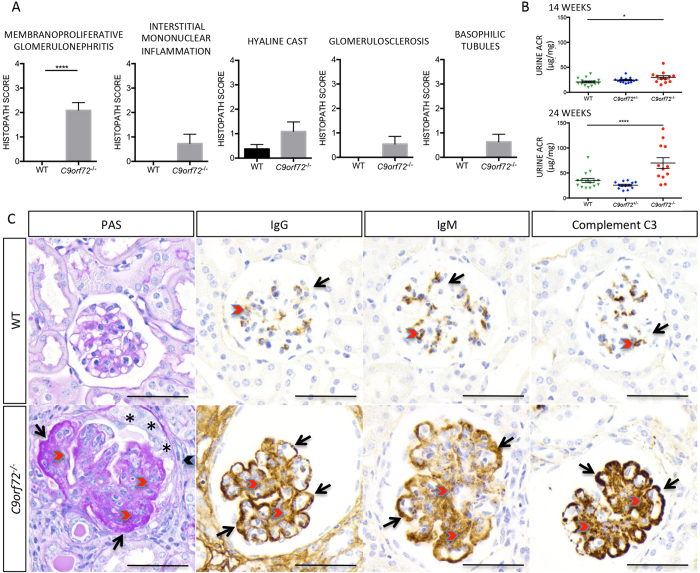
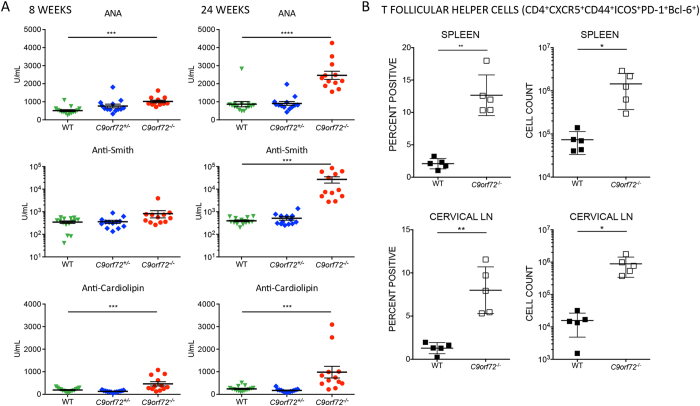
The expansion of a hexanucleotide (GGGGCC) repeat in C9ORF72 is the most common cause of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Both the function of C9ORF72 and the mechanism by which the repeat expansion drives neuropathology are unknown. To examine whether C9ORF72 haploinsufficiency induces neurological disease, we created a C9orf72-deficient mouse line. Null mice developed a robust immune phenotype characterized by myeloid expansion, T cell activation, and increased plasma cells. Mice also presented with elevated autoantibodies and evidence of immune-mediated glomerulonephropathy. Collectively, our data suggest that C9orf72 regulates immune homeostasis and an autoimmune response reminiscent of systemic lupus erythematosus (SLE) occurs in its absence. We further imply that haploinsufficiency is unlikely to be the causative factor in C9ALS/FTD pathology.
Figures
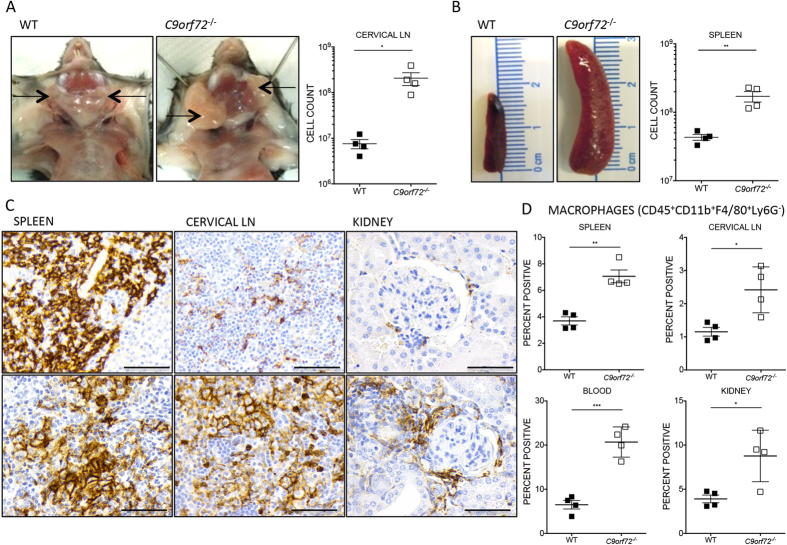
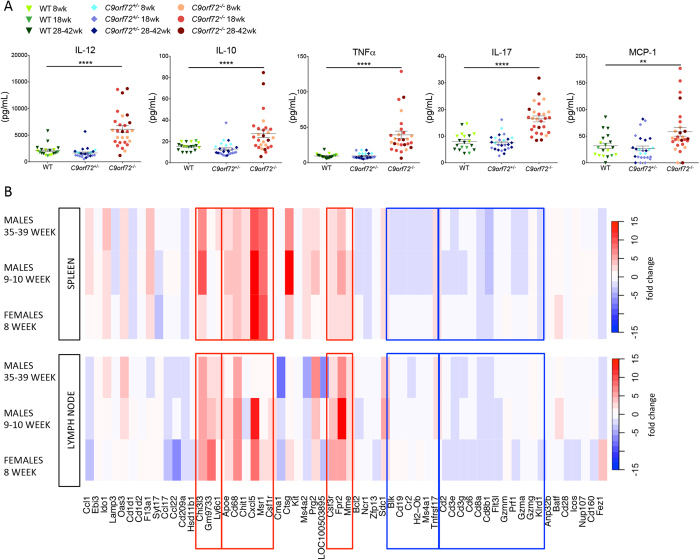
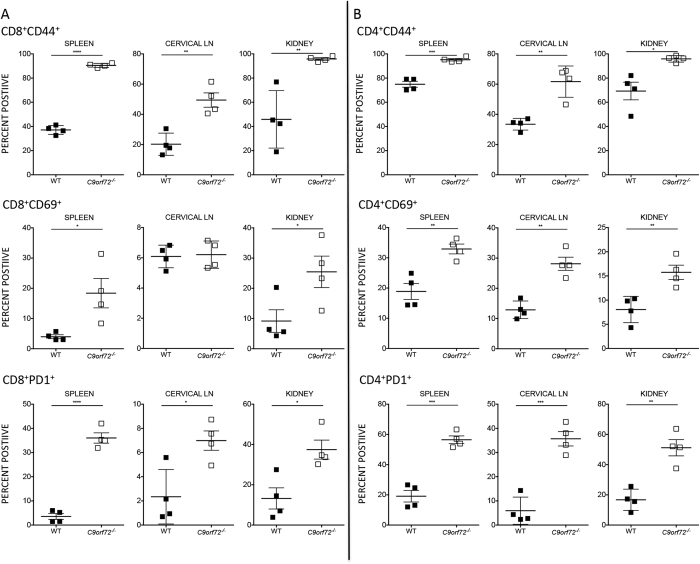
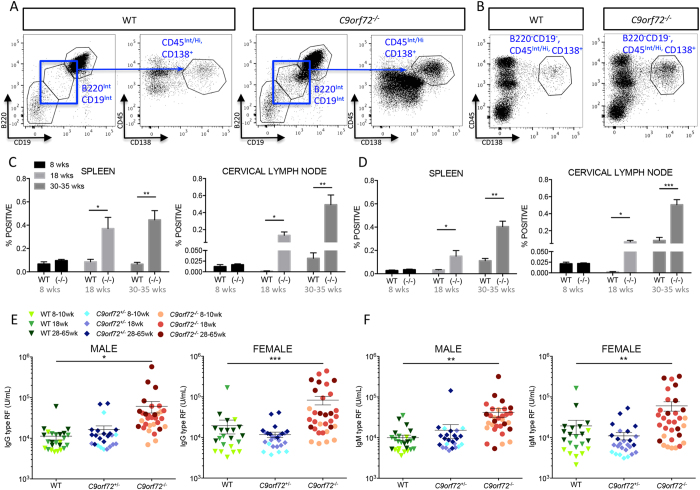
Comment in
-
Full ablation of C9orf72 in mice causes immune system-related pathology and neoplastic events but no motor neuron defects.Acta Neuropathol. 2016 Jul;132(1):145-7. doi: 10.1007/s00401-016-1581-x. Epub 2016 May 20. Acta Neuropathol. 2016. PMID: 27206760 Free PMC article. No abstract available.
References
-
- van Swieten J. C. & Grossman M. FTD/ALS families are no longer orphaned: the C9ORF72 story. Neurology 79, 962–964 (2012). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
