

An apple rootstock overexpressing a peach CBF gene alters growth and flowering in the scion but does not impact cold hardiness or dormancy
- PMID: 26981253
- PMCID: PMC4783695
- DOI: 10.1038/hortres.2016.6
An apple rootstock overexpressing a peach CBF gene alters growth and flowering in the scion but does not impact cold hardiness or dormancy
Abstract
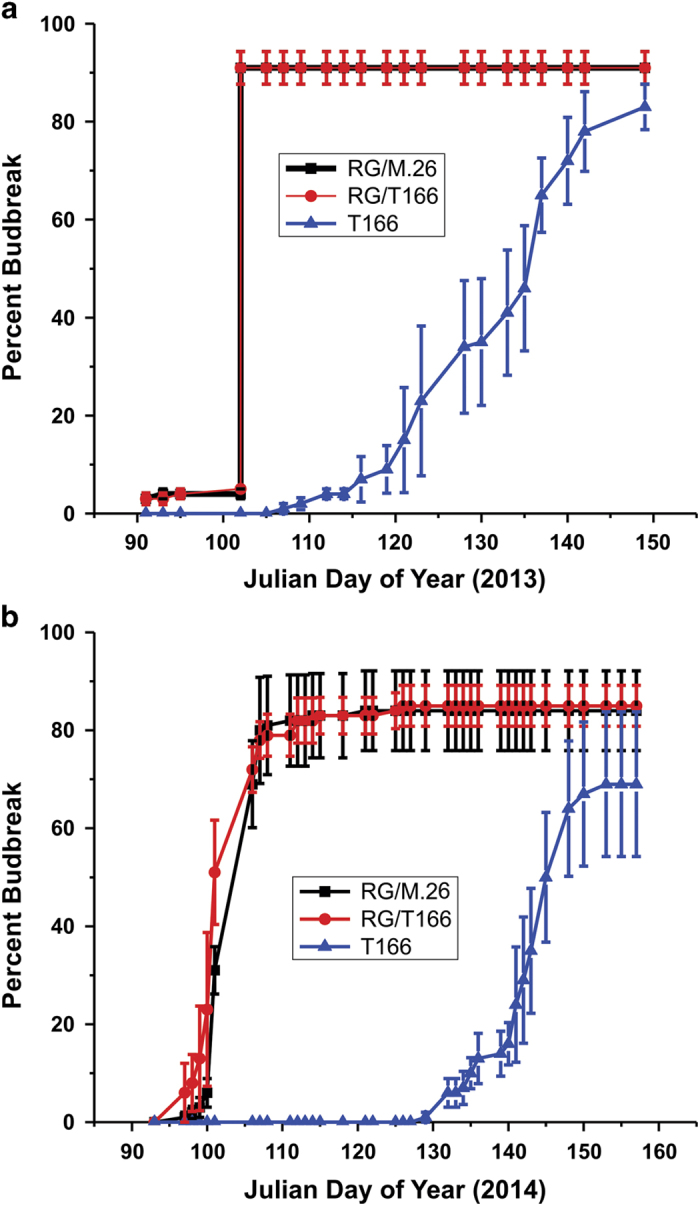
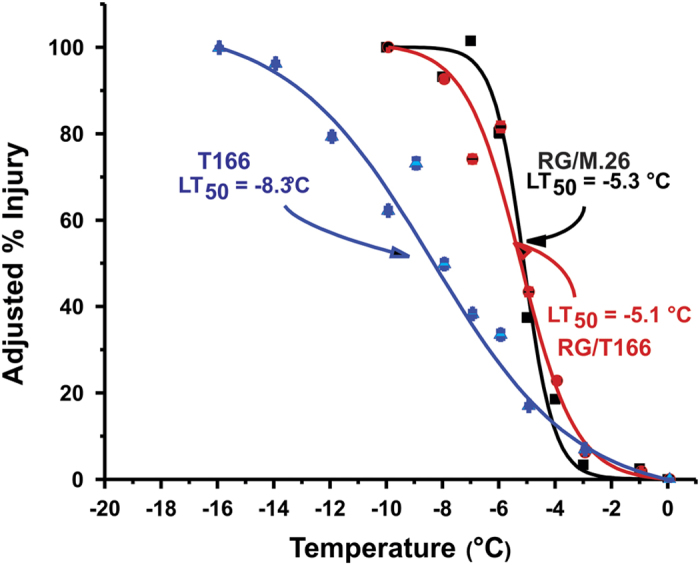
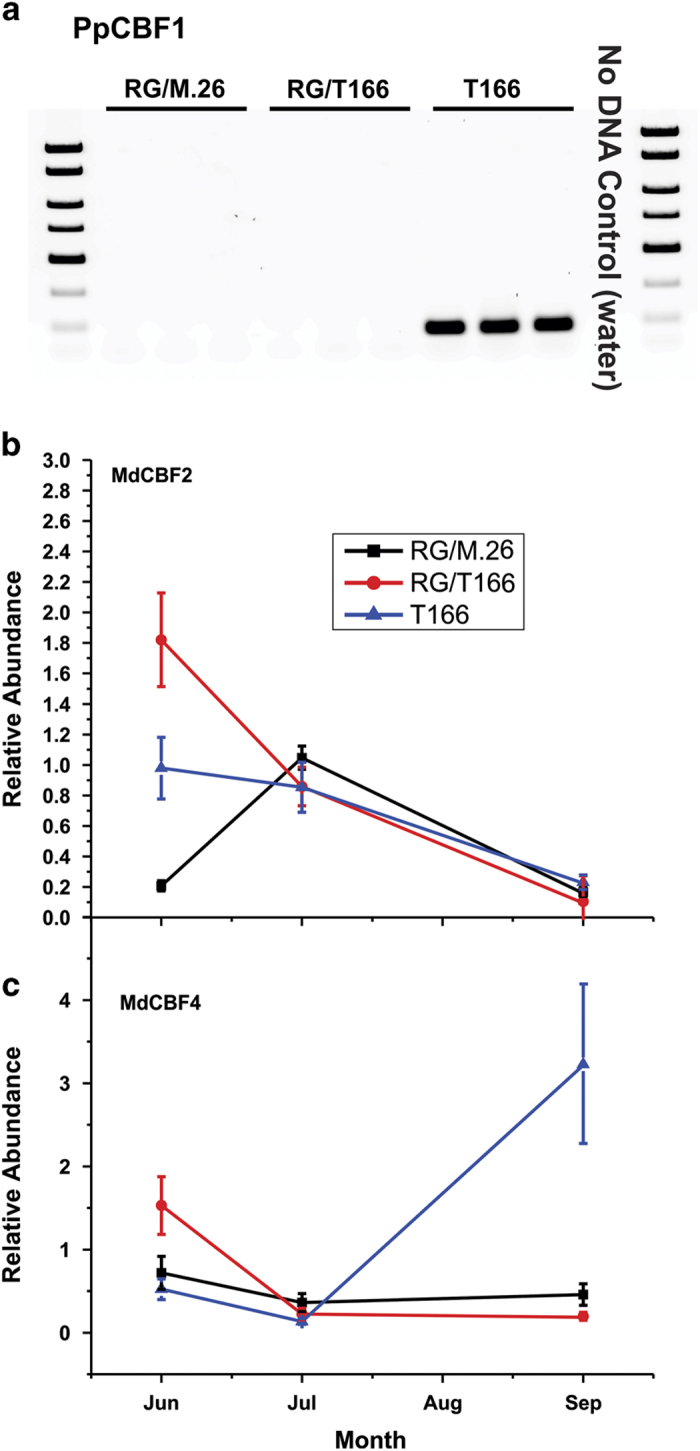
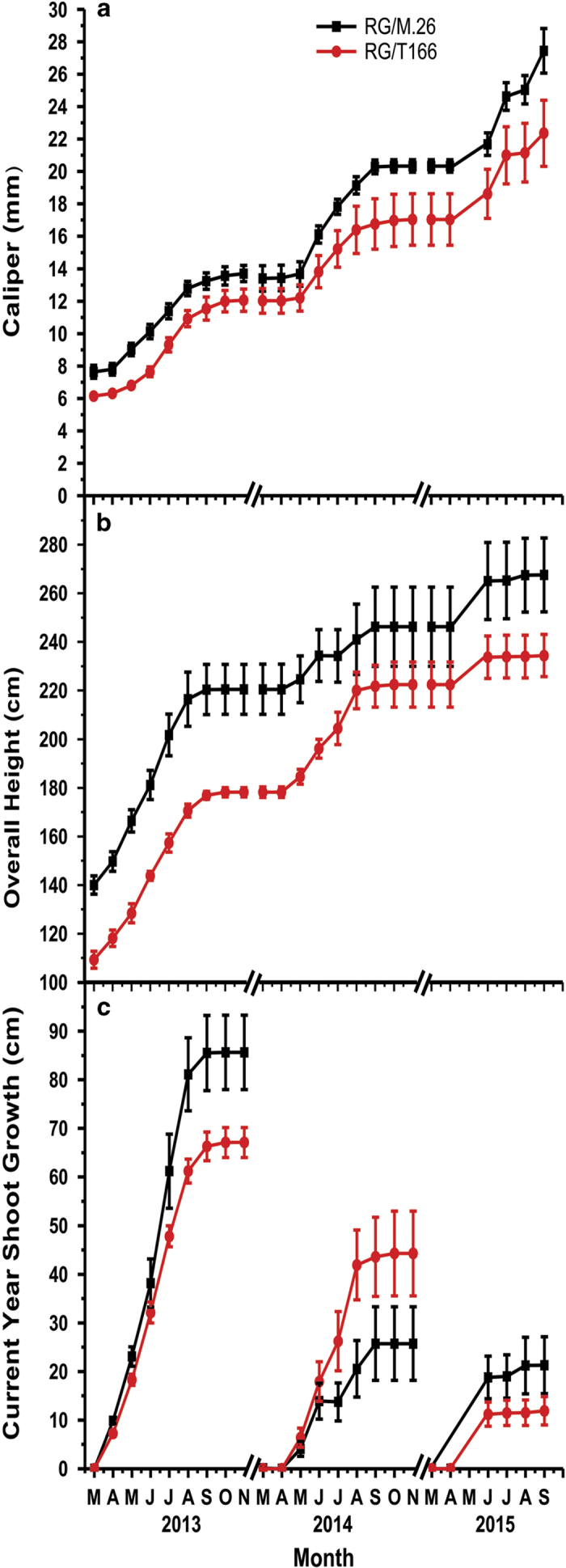
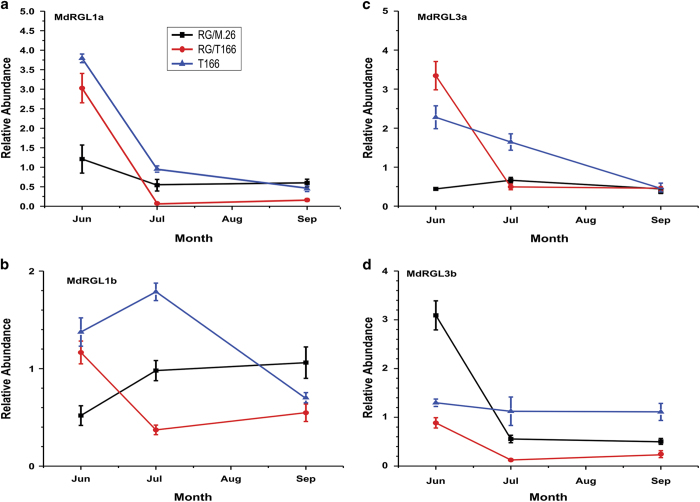
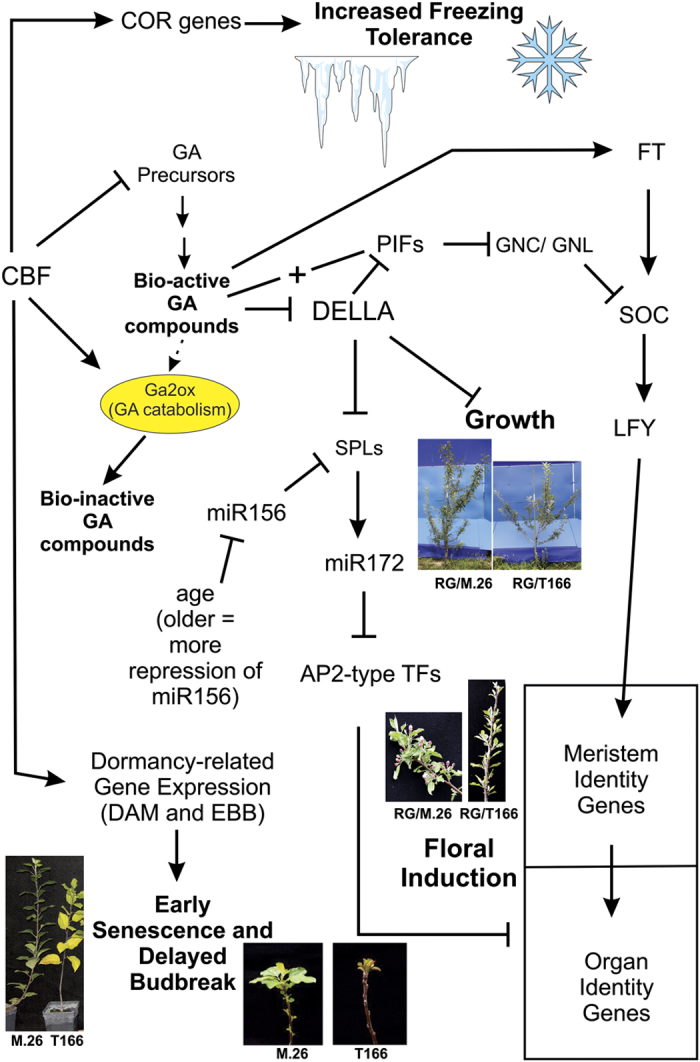
The C-repeat binding factor (CBF) transcription factor is involved in responses to low temperature and water deficit in many plant species. Overexpression of CBF genes leads to enhanced freezing tolerance and growth inhibition in many species. The overexpression of a peach CBF (PpCBF1) gene in a transgenic line of own-rooted apple (Malus×domestica) M.26 rootstock (T166) trees was previously reported to have additional effects on the onset of dormancy and time of spring budbreak. In the current study, the commercial apple cultivar 'Royal Gala' (RG) was grafted onto either non-transgenic M.26 rootstocks (RG/M.26) or transgenic M.26 (T166) rootstocks (RG/T166) and field grown for 3 years. No PpCBF1 transcript was detected in the phloem or cambium of RG scions grafted on T166 rootstocks indicating that no graft transmission of transgene mRNA had occurred. In contrast to own-rooted T166 trees, no impact of PpCBF1 overexpression in T166 rootstocks was observed on the onset of dormancy, budbreak or non-acclimated leaf-cold hardiness in RG/T166 trees. Growth, however, as measured by stem caliper, current-year shoot extension and overall height, was reduced in RG/T166 trees compared with RG/M.26 trees. Although flowering was evident in both RG/T166 and RG/M.26 trees in the second season, the number of trees in flower, the number of shoots bearing flowers, and the number of flower clusters per shoot was significantly higher in RG/M.26 trees than RG/T166 trees in both the second and third year after planting. Elevated levels of RGL (DELLA) gene expression were observed in RG/T166 trees and T166 trees, which may play a role in the reduced growth observed in these tree types. A model is presented indicating how CBF overexpression in a rootstock might influence juvenility and flower abundance in a grafted scion.
Figures


References
-
- Medina J , Catalá R , Salinas J . The CBFs: three arabidopsis transcription factors to cold acclimate. Plant Sci 2011; 180: 3–11. - PubMed
-
- Qin F , Shinozaki K , Yamaguchi-Shinozaki K . Achievements and challenges in understanding plant abiotic stress responses and tolerance. Plant Cell Physiol 2011; 52: 1569–1582. - PubMed
-
- Mizoi J , Shinozaki K , Yamaguchi-Shinozaki K . AP2/ERF transcription factors in plant abiotic stress responses. Biochim Biophys Acta 2012; 1819: 86–96. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
