

Loss of Mitochondrial Function Impairs Lysosomes
- PMID: 26987902
- PMCID: PMC4858975
- DOI: 10.1074/jbc.M115.695825
Loss of Mitochondrial Function Impairs Lysosomes
Abstract
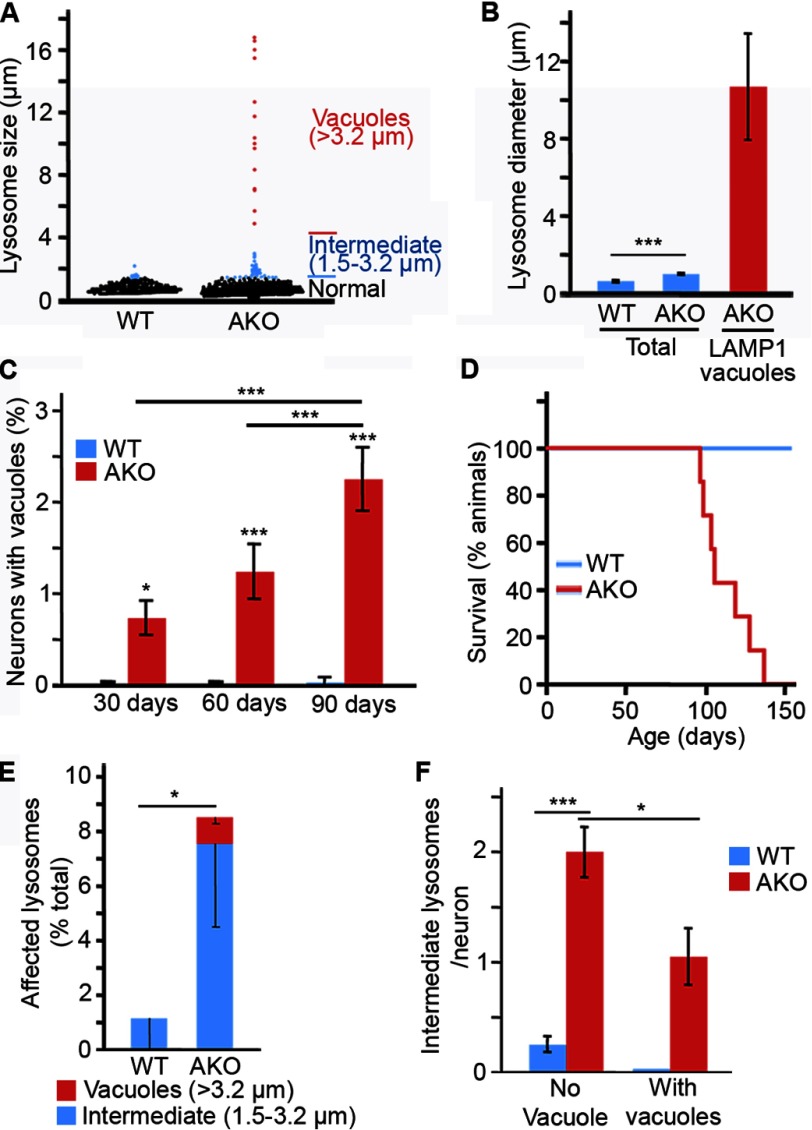
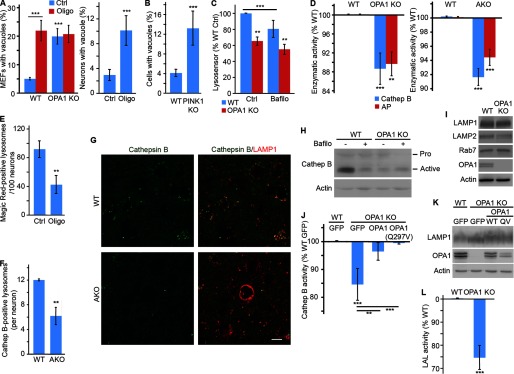
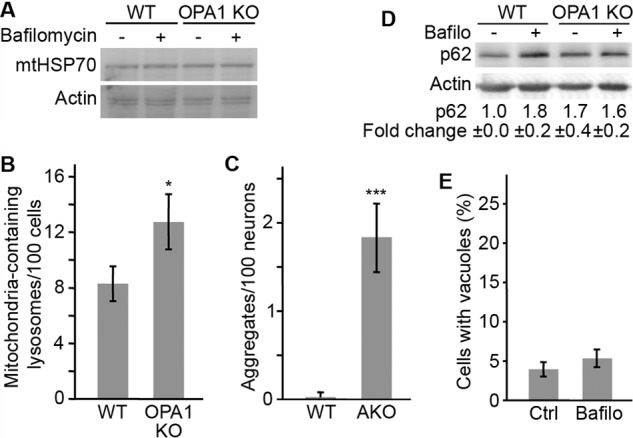
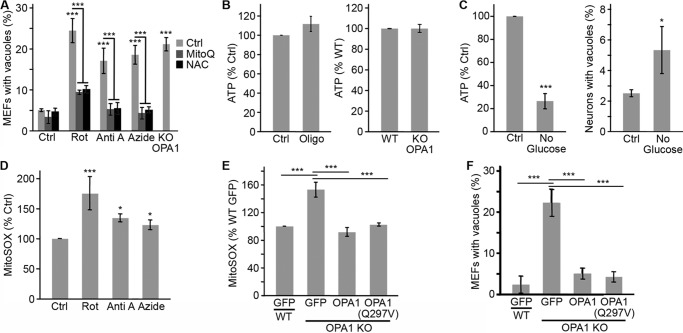
Alterations in mitochondrial function, as observed in neurodegenerative diseases, lead to disrupted energy metabolism and production of damaging reactive oxygen species. Here, we demonstrate that mitochondrial dysfunction also disrupts the structure and function of lysosomes, the main degradation and recycling organelle. Specifically, inhibition of mitochondrial function, following deletion of the mitochondrial protein AIF, OPA1, or PINK1, as well as chemical inhibition of the electron transport chain, impaired lysosomal activity and caused the appearance of large lysosomal vacuoles. Importantly, our results show that lysosomal impairment is dependent on reactive oxygen species. Given that alterations in both mitochondrial function and lysosomal activity are key features of neurodegenerative diseases, this work provides important insights into the etiology of neurodegenerative diseases.
Keywords: Parkin; lysosome; mitochondria; neurodegenerative disease; reactive oxygen species (ROS).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures





References
-
- Ben-Gedalya T., and Cohen E. (2012) Quality control compartments coming of age. Traffic 13, 635–642 - PubMed
-
- Tai H. C., and Schuman E. M. (2008) Ubiquitin, the proteasome and protein degradation in neuronal function and dysfunction. Nat. Rev. Neurosci. 9, 826–838 - PubMed
-
- Patten D. A., Germain M., Kelly M. A., and Slack R. S. (2010) Reactive oxygen species: stuck in the middle of neurodegeneration. J. Alzheimers Dis. 20, S357–S367 - PubMed
-
- Mizushima N., and Komatsu M. (2011) Autophagy: renovation of cells and tissues. Cell 147, 728–741 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
