

Comparative analysis of routes of immunization of a live porcine reproductive and respiratory syndrome virus (PRRSV) vaccine in a heterologous virus challenge study
- PMID: 26988085
- PMCID: PMC4797253
- DOI: 10.1186/s13567-016-0331-3
Comparative analysis of routes of immunization of a live porcine reproductive and respiratory syndrome virus (PRRSV) vaccine in a heterologous virus challenge study
Abstract
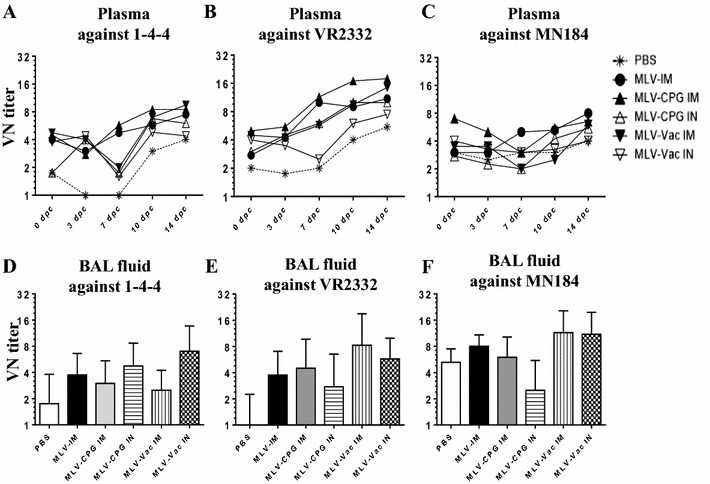
Porcine reproductive and respiratory syndrome (PRRS) is caused by PRRS virus (PRRSV), which infects primarily the respiratory tract of pigs. Thus intranasal (IN) delivery of a potent vaccine-adjuvant formulation is promising. In this study, PRRS-MLV (VR2332) was coadministered ± an adjuvant Mycobacterium vaccae whole cell lysate or CpG ODN through intramuscular (IM) or IN route as a mist, and challenged with a heterologous PRRSV 1-4-4 IN at 42 days post-vaccination (dpv). At 14 and 26 dpv, vaccine viral RNA copies were one log greater in the plasma of PRRS-MLV IM compared to IN vaccinated pigs, and the infectious replicating vaccine virus was detected only in the IM group. In PRRS-MLV ± adjuvant IM vaccinated pigs, reduced viral RNA load and absence of the replicating challenged virus was observed at 7, 10 and 14 days post-challenge (dpc). At 14 dpc, in BAL fluid ≥ 5 log viral RNA copies were detected in all the pig groups, but the replicating challenged virus was undetectable only in IM groups. Immunologically, virus neutralizing antibody titers in the plasma of IM (but not IN) vaccine groups was ≥ 8 against the vaccine and challenged viruses. At 26 dpv, PRRS-MLV IM (without adjuvant) received pigs had significantly increased population of CD4 and CD8 T cells in PBMC. At 14 dpc, relatively increased population of IFN-γ(+) total lymphocytes, NK, CD4, CD8 and γδ T cells were observed in the MLV-IM group. In conclusion, PRRS-MLV IM vaccination induced the virus specific T cell response in pigs, but still it is required to improve its cross-protective efficacy.
Figures




References
-
- Neumann EJ, Kliebenstein JB, Johnson CD, Mabry JW, Bush EJ, Seitzinger AH, Green AL, Zimmerman JJ. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J Am Vet Med Assoc. 2005;227:385–392. doi: 10.2460/javma.2005.227.385. - DOI - PubMed
-
- Holtkamp DJ, Kliebenstein JB, Neumann EJ, Zimmerman JJ, Rotto HF, Yoder TK, Wang C, Yeske PE, Mowrer CL, Haley CA. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J Swine Health Prod. 2013;21:72–84.
-
- Collins JE, Benfield DA, Christianson WT, Harris L, Hennings JC, Shaw DP, Goyal SM, McCullough S, Morrison RB, Joo HS, Gorcyca D, Chladek D. Isolation of swine infertility and respiratory syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic pigs. J Vet Diagn Invest. 1992;4:117–126. doi: 10.1177/104063879200400201. - DOI - PubMed
-
- Wensvoort G, Terpstra C, Pol JM, ter Laak EA, Bloemraad M, de Kluyver EP, Kragten C, van Buiten L, den Besten A, Wagenaar F, Broekhuijsen JM, Moonen PL, Zetstra T, de Boer EA, Tibben HJ, de Jong MF, van’t Veld P, Greenland GJ, van Gennep JA, Voets MT, Verheijden JH, Braamskamp J. Mystery swine disease in The Netherlands: the isolation of lelystad virus. Vet Q. 1991;13:121–130. doi: 10.1080/01652176.1991.9694296. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
