

Review
doi: 10.1101/cshperspect.a018838.
Diversity of Neural Precursors in the Adult Mammalian Brain
Affiliations
- PMID: 26988967
- PMCID: PMC4817799
- DOI: 10.1101/cshperspect.a018838
Item in Clipboard
Review
Diversity of Neural Precursors in the Adult Mammalian Brain
Cold Spring Harb Perspect Biol.
.
Abstract
Aided by advances in technology, recent studies of neural precursor identity and regulation have revealed various cell types as contributors to ongoing cell genesis in the adult mammalian brain. Here, we use stem-cell biology as a framework to highlight the diversity of adult neural precursor populations and emphasize their hierarchy, organization, and plasticity under physiological and pathological conditions.
Copyright © 2016 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
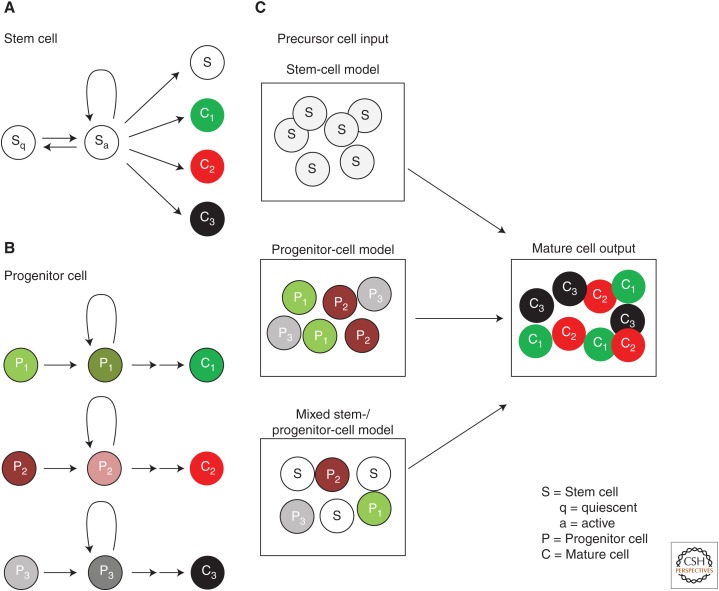
Models of generating cell diversity in the adult tissues. (A,B) Definitions of stem and progenitor cells. In A, quiescent stem cells (Sq) become active stem cells (Sa) that proliferate to generate different types of specialized cells (C1, C2, C3) and new stem cells (S). The active stem cell can return to quiescence and remain quiescent over long periods of time. In B, lineage-restricted progenitor cells lacking self-renewal capacity (P1, P2, P3) each give rise to distinct populations of specialized cells (C1, C2, C3). (C) Generation of specialized cells in a tissue could be explained by three models. (1) The stem-cell model, in which multipotent stem cells give rise to all the specialized cells in the tissue. (2) The progenitor cell model, in which diverse, lineage-restricted progenitor cells give rise to different cell types in the tissue. (3) A hybrid model, in which a mixture of stem cells and lineage-restricted progenitor cells generate specialized cells of the adult tissue.
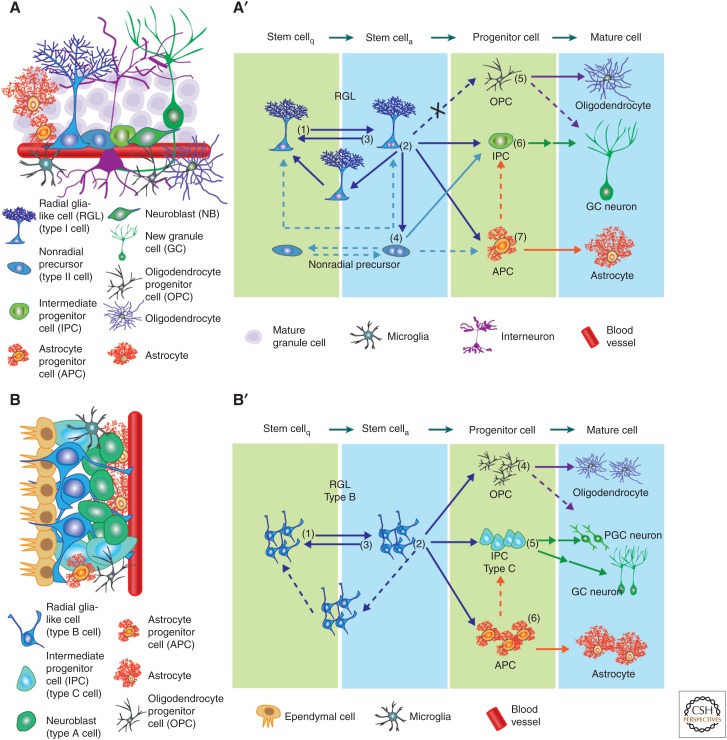
The stem-cell niche and lineage hierarchy among stem cells and their progeny. (A) Schematic illustration of the cell types present in the neurogenic niche of subgranular zone (SGZ) in the dentate gyrus (DG). Shown in A′ is a model of the neural genesis in the adult SGZ under basal conditions. On the single-cell level, quiescent radial glia-like cells (RGLs) can become active and enter cell cycle (1). The active RGLs can generate an intermediate progenitor cell (IPC), an astrocyte progenitor cell (APC), or generate an additional RGL (2), which can become either quiescent or active. The active RGLs can remain in a proliferative state or return to quiescence (3). Nonradial, nestin+ cells produce IPCs and may cycle between quiescent and active states (4). Oligodendrocyte progenitor cells (OPCs) can enter cell cycle and generate mature oligodendrocytes (5). IPCs generate excitatory granule cell (GC) neurons (6). Newborn APCs can generate mature astrocytes (7). (B) A schematic illustration of the cell types present in the neurogenic niche of the subventricular zone (SVZ). Shown in B′ is a model of the neural genesis in the adult SVZ under basal conditions. On the population level (indicated by more than one cell), quiescent RGLs (type B cells) can become active and enter cell cycle (1). Progeny of active type B cells can give rise to IPCs (type C cells), APCs, and OPCs. Whether the RGLs directly make these fate choices remains unknown. RGLs may also generate additional type B cells, which could be either active or quiescent. Active type B cells can return to quiescence (3). OPCs can enter cell cycle and generate mature oligodendrocytes (4). IPCs generate classes of inhibitory neurons including periglomerular cell (PGC) and GC neurons, although it remains unclear whether the fate decisions occur within RGLs or IPCs (5). Newborn APCs can generate mature astrocytes (6). Arrows indicate direct cell generation. Dotted arrows represent potential choices requiring further experimental evidence. Double arrows represent multistep cell generation. Arrows with an X represent choices not experimentally observed.
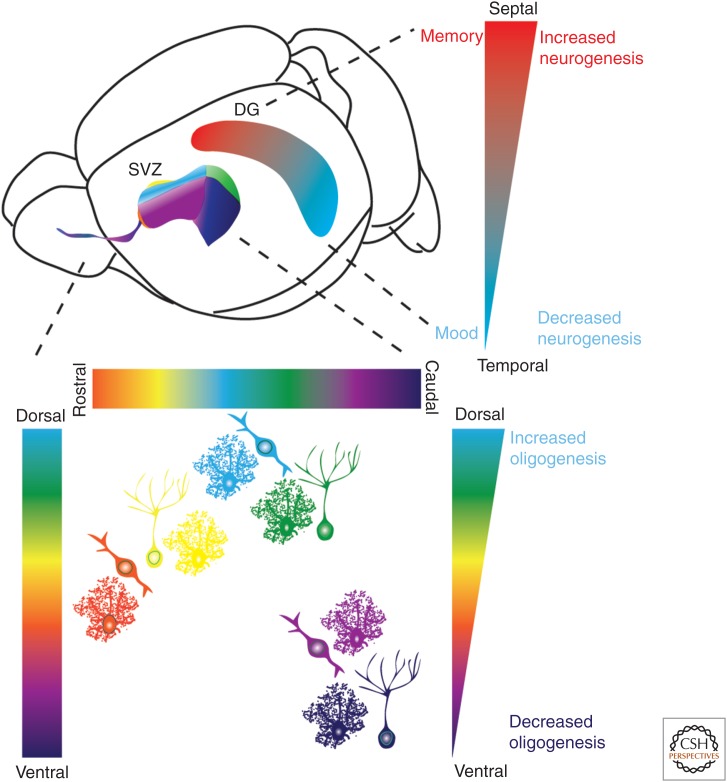
Regionalization of adult neural genesis in classical neurogenic zones. A schematic illustration of the mouse brain indicating the location of SVZ/RMS (subventricular zone/rostral migratory stream) and subgranular zone (SGZ) neurogenic niches. The different colors denote spatial heterogeneity of neural precursor cells. Precursor cells within the SVZ generate different interneuron subtypes, granule cells (GCs) or pergriglomerular (periglomerular cells [PGCs]; see Fig. 2). Studies have further divided these cell types in which precursor cells originating from the ventral SVZ generate deep granule cell (purple) and calbindin-expressing periglomerular cells (magenta). Precursor cells from the dorsal SVZ on the other hand, generate superficial granule cells (green) and TH+ periglomerular cells (light blue), whereas precursor cells in the medial parts of the ventricular wall generate calretinin-expressing superficial granule cells (yellow) and periglomerular cells (orange). Contrary to spatial heterogeneity and migration of long distances of neuron subtypes, astrocytes are generated throughout the SVZ and migrate more locally. Oligodendrogenesis is higher in the dorsal SVZ compared with the lateral and ventral SVZ. As schematically illustrated, the septal part of the dentate gyrus (DG) is associated with memory, has higher neurogenesis within the SGZ compared with the temporal part, which in turn has lower neurogenesis and is associated with mood regulation.
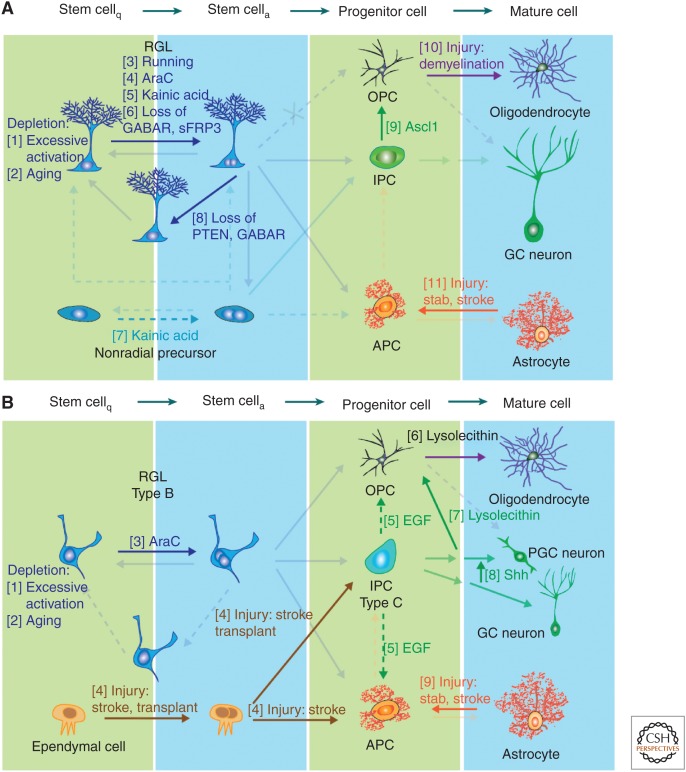
Regulation of neural precursor plasticity within the classical neurogenic zones. Schematic illustration of example factors and manipulations known to regulate cell genesis in the adult subgranular zone (SGZ) (A) and subventricular zone (SVZ) (B). Numbers denote examples known to affect lineage decisions at the stage indicated in the figure. (A) Stem-cell loss occurs when their proliferation is highly induced, such as through Notch and FoxO deletion (1) (Paik et al. 2009; Renault et al. 2009; Ehm et al. 2010; Imayoshi et al. 2010), or in aged mice (2) (Kuhn et al. 1996; Encinas et al. 2011; Villeda et al. 2011). Mobilization of quiescent radial glia-like cells (RGLs) occurs during voluntary running (3) (Kempermann et al. 1997; van Praag et al. 1999); brain injury, such as injection of the antimitotic drug Ara-C (Seri et al. 2001) (4) or seizure-inducing Kainic acid (5) (Steiner et al. 2008; Jiruska et al. 2013). Molecular inhibitors of RGL activation include SFRP3 and GABA signaling (6) (Song et al. 2012; Jang et al. 2013). Kainic acid-induced seizures activate nonradial progenitor cells (7) (Lugert et al. 2010). Increasing Akt signaling or decreasing tonic GABA signaling alters the division mode of RGLs, fostering the symmetric fate (8) (Bonaguidi et al. 2011; Song et al. 2012). Ectopic expression of Ascl1 changes the fate of intermediate progenitor cells (IPCs) to generate oligodendrocyte progenitor cells (OPCs) (9) (Jessberger et al. 2008) and demyelination injury induces OPC proliferation (10) (Nait-Oumesmar et al. 1999; Menn et al. 2006; Hughes et al. 2013). Stab wound, stroke and ischemic injuries activate astrocytes into reactive astroglia (11) (reviewed in Robel et al. 2011). (B) In the SVZ excessive activation (1) (Paik et al. 2009; Renault et al. 2009; Ehm et al. 2010; Imayoshi et al. 2010) and aging (2) (Kuhn et al. 1996; Molofsky et al. 2006; Villeda et al. 2011) leads to stem-cell loss. Ara-C promotes RGL cell-cycle entry (3) (Doetsch et al. 1999) and stroke injury activates the normally quiescent ependymal cells (4) (Johansson et al. 1999; Coskun et al. 2008; Carlen et al. 2009). Infusion of EGF increases production of astroglia and OPCs while reducing proliferation of IPCs (5) (Craig et al. 1996; Kuhn et al. 1997). Demyelination injury increases OPC proliferation (6) and doublecortin (DCX)+ neural progenitors to swich fate into OPCs (7) (Nait-Oumesmar et al. 1999; Menn et al. 2006; Jablonska et al. 2010; Hughes et al. 2013). Manipulation of the Sonic hedgehog (SHH) signaling pathway can change the fate of a subset of neural progenitors from granule cell (GC) neurons to periglomerular cell (PGC) neurons (8) (Ihrie et al. 2011). Stab wound, stroke, and ischemic injuries activate astrocytes into reactive astroglia (9) (reviewed in Robel et al. 2011).
References
-
- Ahn S, Joyner AL. 2005. In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature 437: 894–897. - PubMed
-
- Altman J, Bayer SA. 1990. Migration and distribution of two populations of hippocampal granule cell precursors during the perinatal and postnatal periods. J Comp Neurol 301: 365–381. - PubMed
-
- Alvarez-Buylla A, Kohwi M, Nguyen TM, Merkle FT. 2008. The heterogeneity of adult neural stem cells and the emerging complexity of their niche. Cold Spring Harb Symp Quant Biol 73: 357–365. - PubMed
-
- Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O. 2002. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat Med 8: 963–970. - PubMed
-
- Bardehle S, Kruger M, Buggenthin F, Schwausch J, Ninkovic J, Clevers H, Snippert HJ, Theis FJ, Meyer-Luehmann M, Bechmann I, et al. 2013. Live imaging of astrocyte responses to acute injury reveals selective juxtavascular proliferation. Nat Neurosci 16: 580–586. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources