

Repurposing Treprostinil for Enhancing Hematopoietic Progenitor Cell Transplantation
- PMID: 26989084
- PMCID: PMC4885501
- DOI: 10.1124/mol.116.103267
Repurposing Treprostinil for Enhancing Hematopoietic Progenitor Cell Transplantation
Erratum in
-
Corrigendum to "Repurposing Treprostinil for Enhancing Hematopoietic Progenitor Cell Transplantation".Mol Pharmacol. 2025 Mar;107(3):100025. doi: 10.1016/j.molpha.2025.100025. Epub 2025 Feb 21. Mol Pharmacol. 2025. PMID: 39986243 Free PMC article. No abstract available.
Abstract
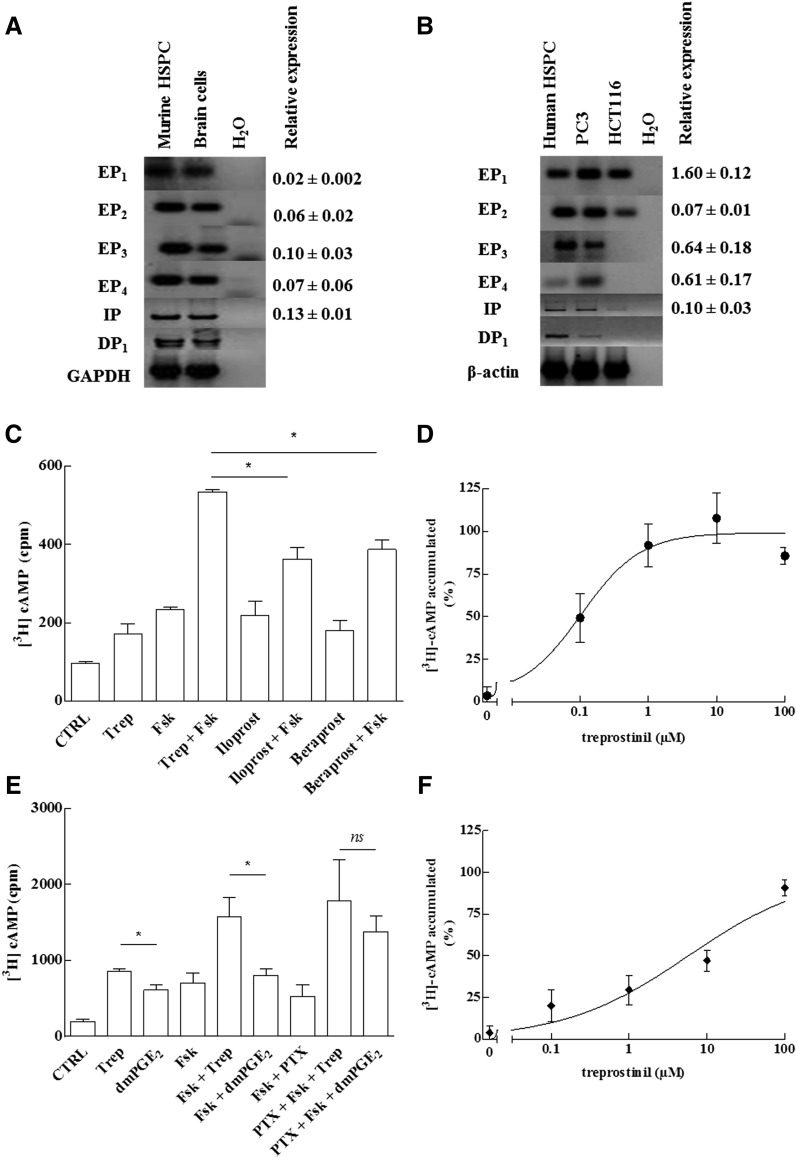
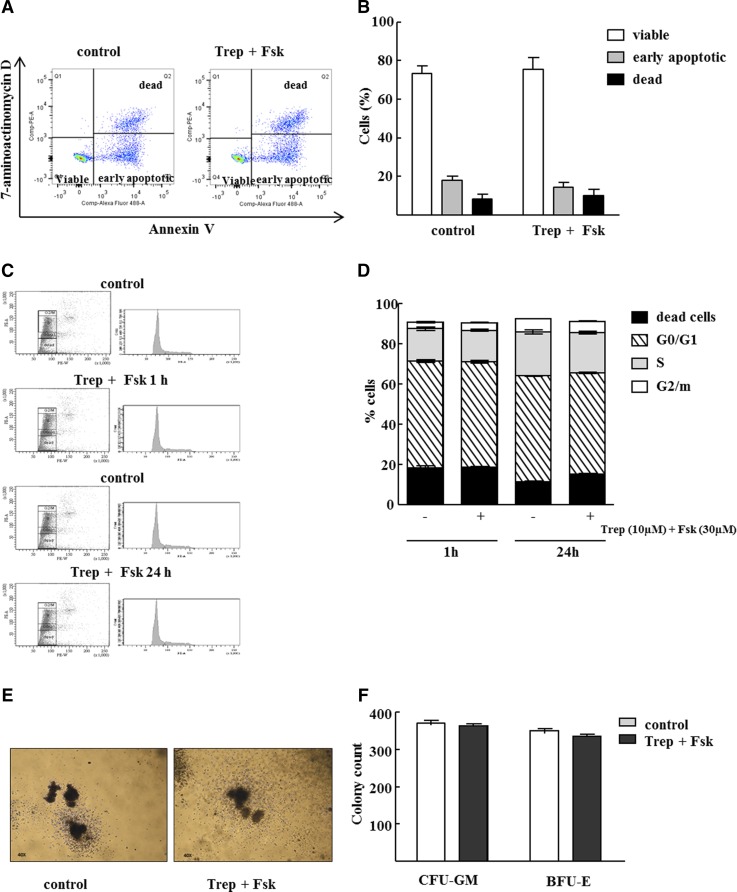
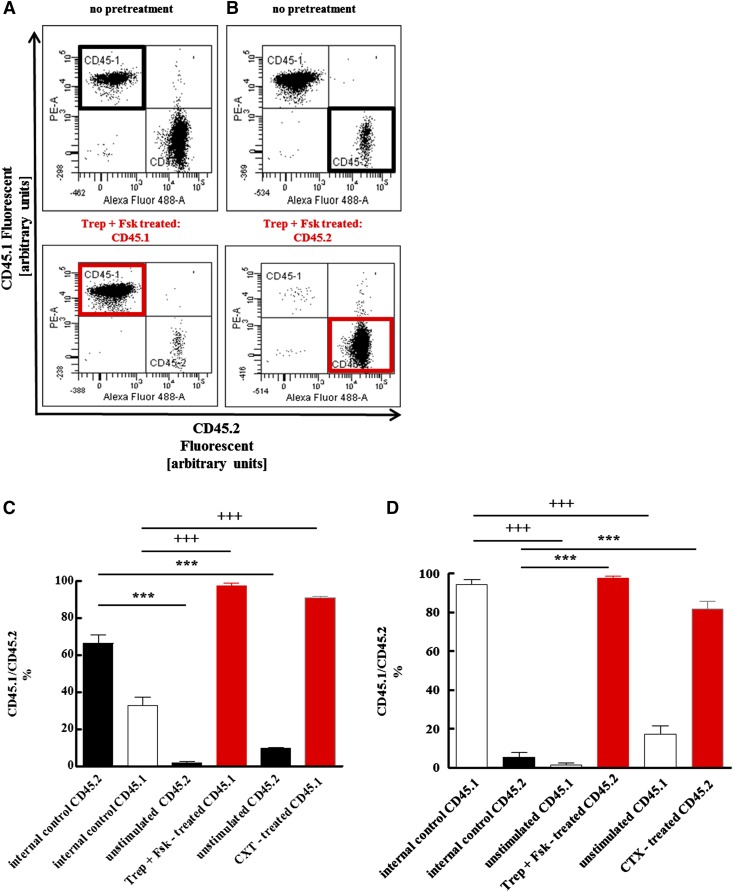
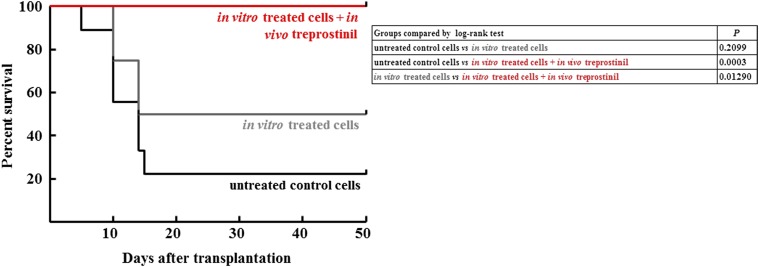
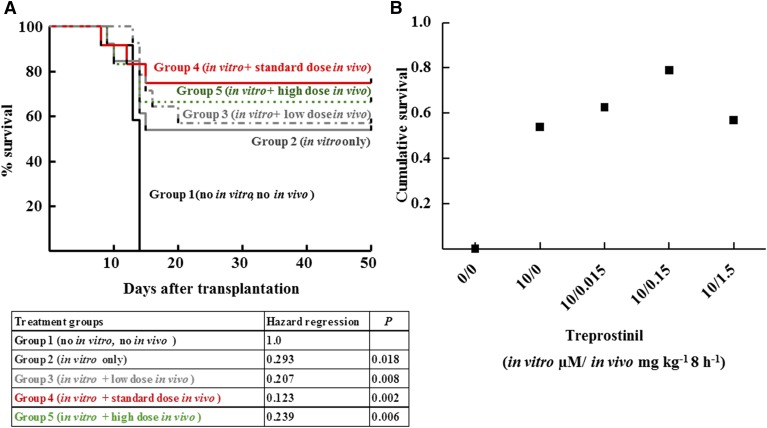
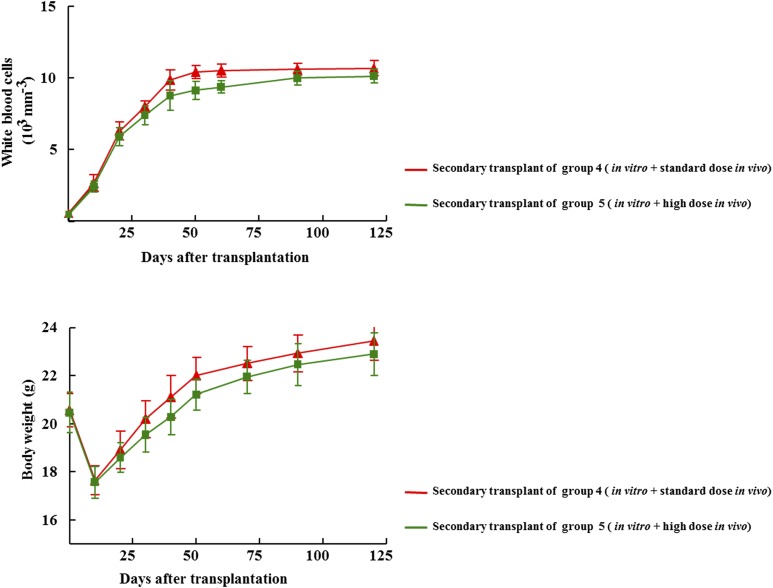
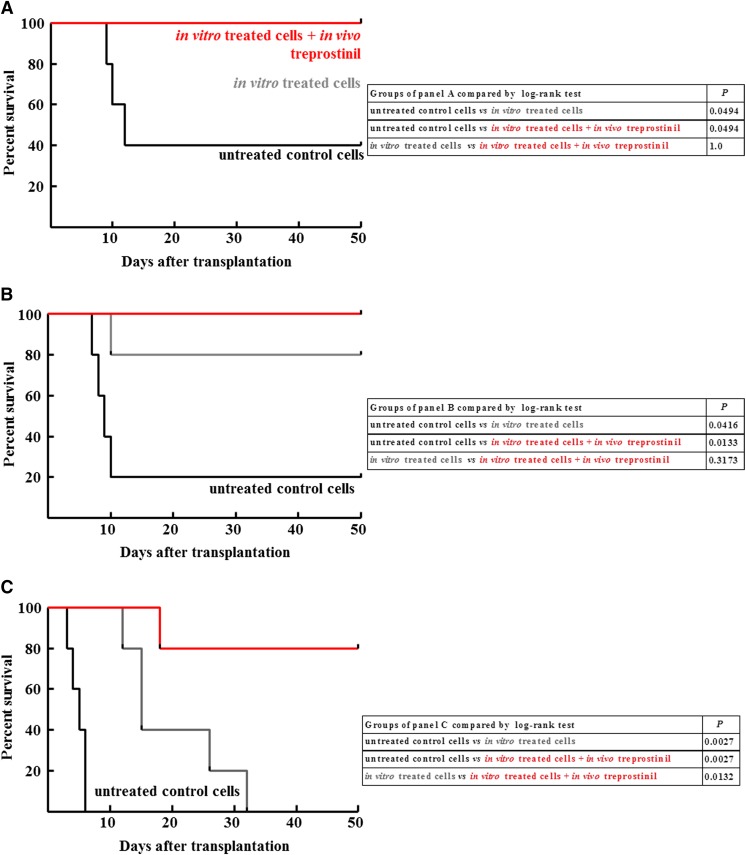
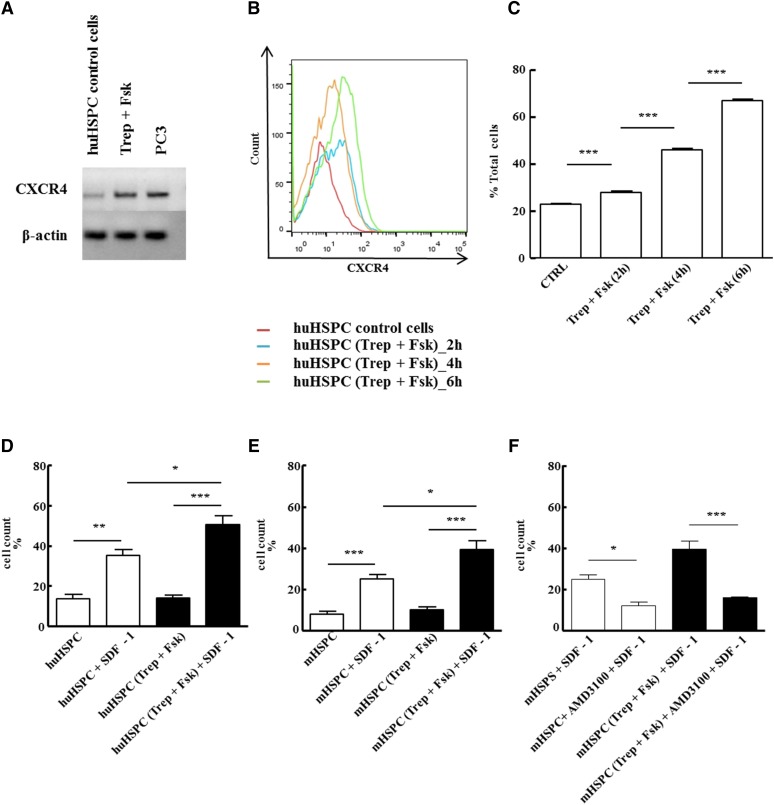
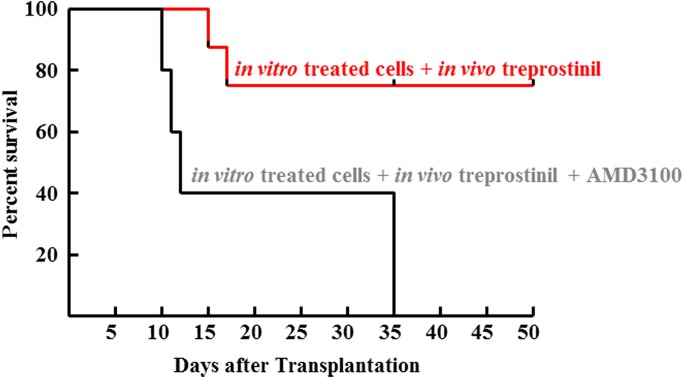
Activation of Gs-coupled receptors enhances engraftment of hematopoietic stem and progenitor cells (HSPCs). We tested the hypothesis that treprostinil, a prostacyclin analog approved for the treatment of pulmonary hypertension, can be repurposed to improve hematopoietic stem cell transplantation. Murine and human HSPCs were isolated from bone marrow and umbilical cord blood, respectively. Prostanoid receptor agonists and the combination thereof with forskolin were tested for their capacity to stimulate [(3)H]cAMP accumulation in HSPCs. Three independent approaches were employed to verify the ability of agonist-activated HSPCs to reconstitute the bone marrow in lethally irradiated recipient mice. The underlying mechanism was explored in cellular migration assays and by blocking C-X-C motif chemokine receptor 4 (CXCR4). Among several prostanoid agonists tested in combination with forskolin, treprostinil was most efficacious in raising intracellular cAMP levels in murine and human HPSCs. Injection of murine and human HSPCs, which had been pretreated with treprostinil and forskolin, enhanced survival of lethally irradiated recipient mice. Survival was further improved if recipient mice were subcutaneously administered treprostinil (0.15 mg kg(-1) 8 h(-1)) for 10 days. This regimen also reduced the number of HSPCs required to rescue lethally irradiated mice. Enhanced survival of recipient mice was causally related to treprostinil-enhanced CXCR4-dependent migration of HSPCs. Treprostinil stimulates the engraftment of human and murine hematopoietic stem cells without impairing their capacity for self-renewal. The investigated dose range corresponds to the dose approved for human use. Hence, these findings may be readily translated into a clinical application.
Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics.
Figures
References
-
- Adams GB, Chabner KT, Alley IR, Olson DP, Szczepiorkowski ZM, Poznansky MC, Kos CH, Pollak MR, Brown EM, Scadden DT. (2006) Stem cell engraftment at the endosteal niche is specified by the calcium-sensing receptor. Nature 439:599–603. - PubMed
-
- Aksentijevich I, Flinn I. (2002) Chemotherapy and bone marrow reserve: lessons learned from autologous stem cell transplantation. Cancer Biother Radiopharm 17:399–403. - PubMed
-
- Aronoff DM, Peres CM, Serezani CH, Ballinger MN, Carstens JK, Coleman N, Moore BB, Peebles RS, Faccioli LH, Peters-Golden M. (2007) Synthetic prostacyclin analogs differentially regulate macrophage function via distinct analog-receptor binding specificities. J Immunol 178:1628–1634. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
