

Integrin signalling regulates YAP and TAZ to control skin homeostasis
- PMID: 26989177
- PMCID: PMC4874484
- DOI: 10.1242/dev.133728
Integrin signalling regulates YAP and TAZ to control skin homeostasis
Abstract
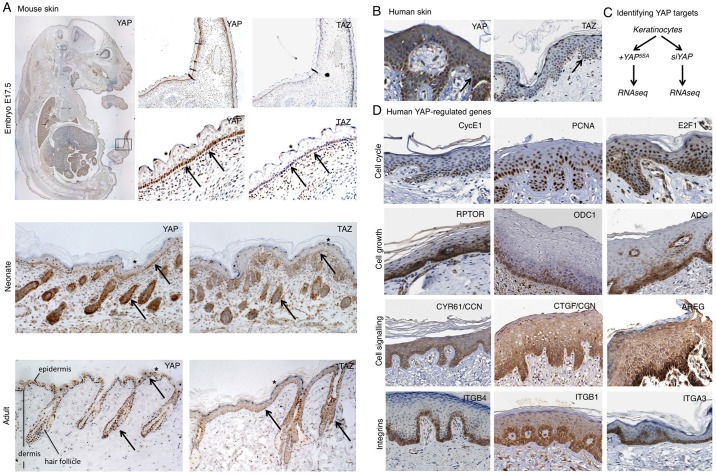
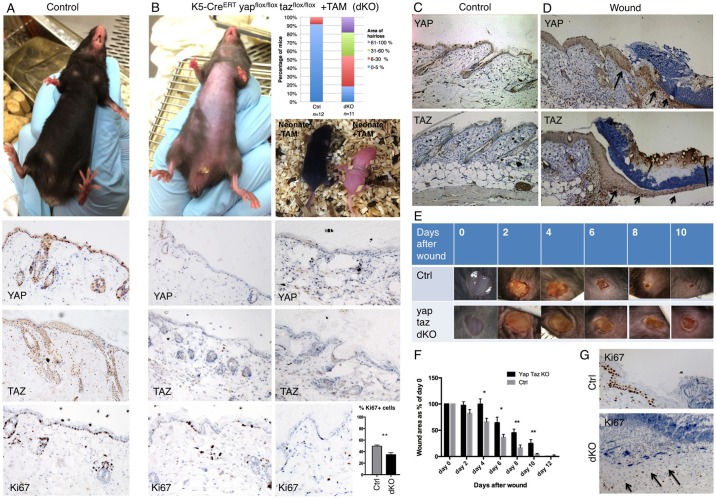
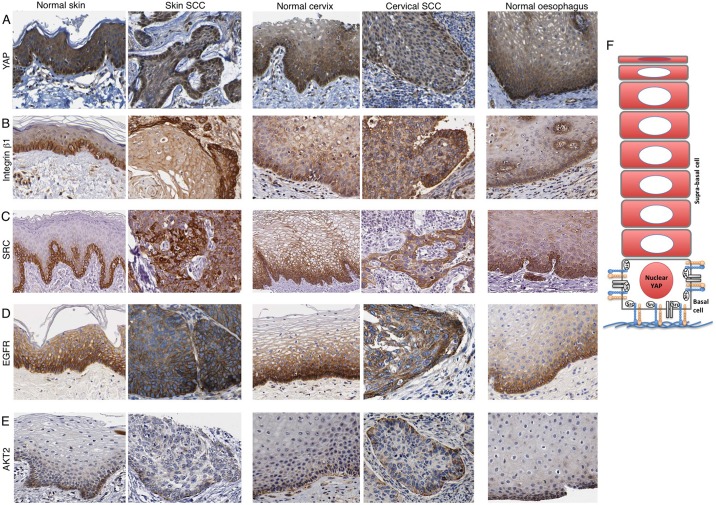
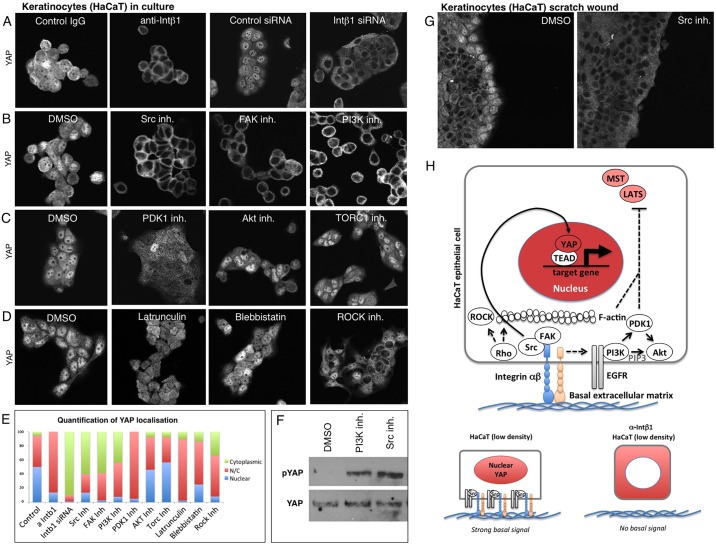
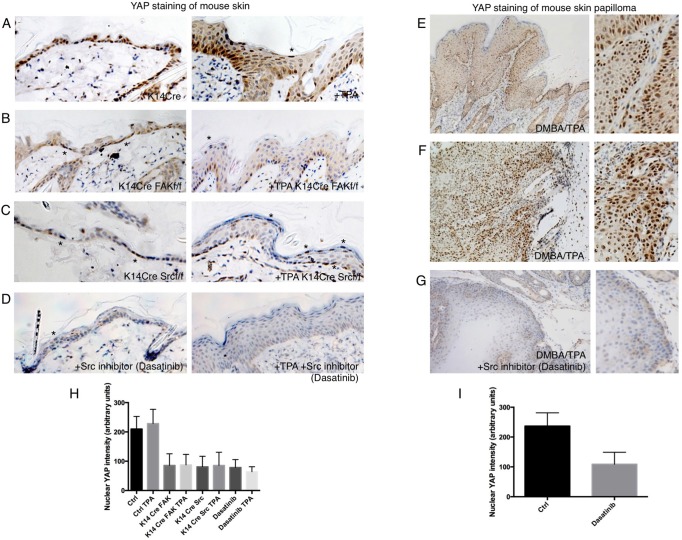
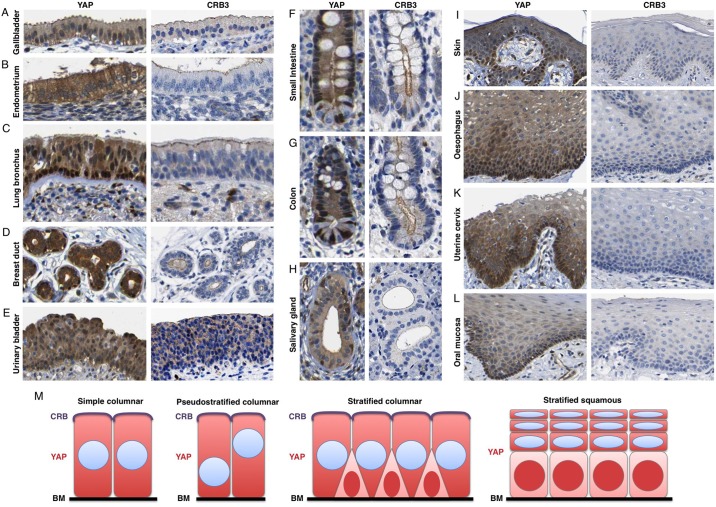
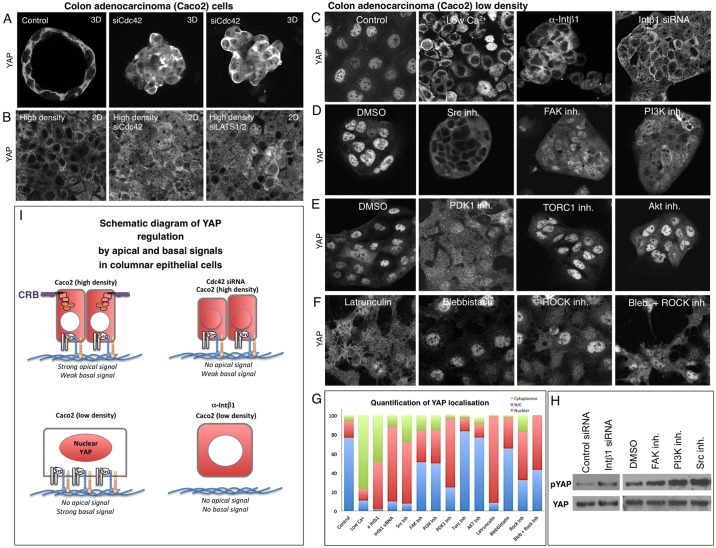
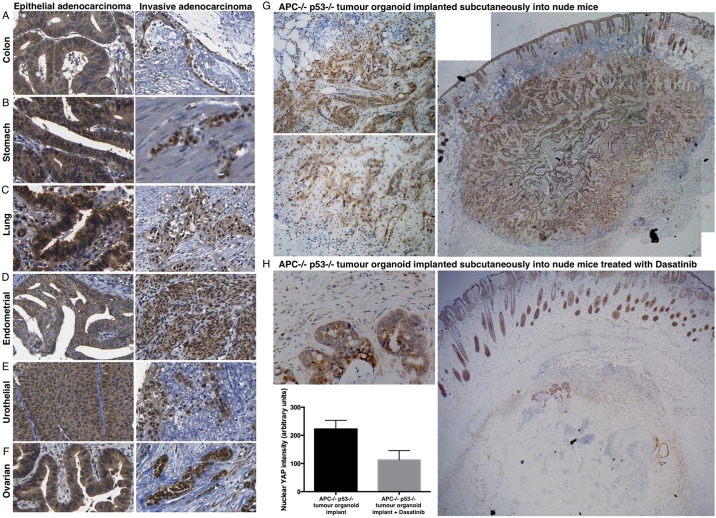
The skin is a squamous epithelium that is continuously renewed by a population of basal layer stem/progenitor cells and can heal wounds. Here, we show that the transcription regulators YAP and TAZ localise to the nucleus in the basal layer of skin and are elevated upon wound healing. Skin-specific deletion of both YAP and TAZ in adult mice slows proliferation of basal layer cells, leads to hair loss and impairs regeneration after wounding. Contact with the basal extracellular matrix and consequent integrin-Src signalling is a key determinant of the nuclear localisation of YAP/TAZ in basal layer cells and in skin tumours. Contact with the basement membrane is lost in differentiating daughter cells, where YAP and TAZ become mostly cytoplasmic. In other types of squamous epithelia and squamous cell carcinomas, a similar control mechanism is present. By contrast, columnar epithelia differentiate an apical domain that recruits CRB3, Merlin (also known as NF2), KIBRA (also known as WWC1) and SAV1 to induce Hippo signalling and retain YAP/TAZ in the cytoplasm despite contact with the basal layer extracellular matrix. When columnar epithelial tumours lose their apical domain and become invasive, YAP/TAZ becomes nuclear and tumour growth becomes sensitive to the Src inhibitor Dasatinib.
Keywords: Hippo pathway; Integrin; Stratified squamous epithelium; TAZ; Yes-associated protein.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
